Paryby
třída obratlovců (Vertebrata) From Wikipedia, the free encyclopedia

Paryby (Chondrichthyes) jsou mořští, vzácně i sladkovodní čelistnatci s chrupavčitou, i když na povrchu zvápenatělou kostrou. Nejznámějšími z nich jsou žraloci a rejnoci (kteří zároveň tvoří dvě podskupiny takzvaných příčnoústých paryb), kromě nich mezi paryby patří ještě nepočetná skupina chimér. Celkem je známo kolem 1200 druhů paryb klasifikovaných asi do 13 řádů. Paryby jsou kromě extrémní redukce kostní tkáně charakteristické také absencí řady krycích lebečních kostí typických pro ostatní čelistnatce (chybí např. skřelové kosti, kosti lebeční klenby, hlavní kosti čelistí, které jsou pak uspořádány odlišně aj.). Jejich tělo je kryto specifickým typem šupin (tzv. plakoidními šupinami se stejnou strukturou, jakou mají zuby). Zuby paryb se zakládají v řadách, které se postupně nasouvají dopředu, kde nahrazují vypadlé zuby. U chimér však došlo během jejich evoluce jednak ke ztrátě plakoidních šupin na většině těla a jejich zuby jsou modifikované v nevyměňované zubní desky.
| Paryby Stratigrafický výskyt: silur až recent | |
|---|---|
_over_havbotn.jpg)  .jpg)  2.jpg) Chiméra podivná (nahoře), žralok obrovský a žralok bílý (uprostřed) a parejnok okatý a siba skvrnitá (dole) | |
| Vědecká klasifikace | |
| Říše | živočichové (Animalia) |
| Kmen | strunatci (Chordata) |
| Podkmen | obratlovci (Vertebrata) |
| Infrakmen | čelistnatci (Gnathostomata) |
| Třída | paryby (Chondrichthyes) Huxley, 1880 |
| Podtřídy | |
| |
| Sesterská skupina | |
| Teleostomi | |
| Některá data mohou pocházet z datové položky. | |
Paryby jsou vybaveny v typickém případě dvěma hřbetními ploutvemi, řitní a ocasní ploutví, která je nejčastěji heterocerkní. Párové končetiny jsou jako u ostatních čelistnatců přítomny ve dvou párech, v tomto případě v podobě prsních a břišních ploutví. Břišní ploutve samců jsou zčásti modifikovány v pářicí orgány (tzv. pterygopody), pomocí nichž probíhá vnitřní oplození. Paryby kladou velká vejce nebo jsou vejcoživorodé, případně rodí živá mláďata vyživovaná různým způsobem v děloze matky. Chybí jim plynový měchýř, jehož roli v nadlehčování těla zastupují játra bohatá tuky. Ve střevě paryb je vyvinuta spirální řasa, která zvětšuje povrch pro vstřebávání živin. Paryby se živí rozmanitou potravou. Mohou lovit různorodou kořist (ryby a další vodní obratlovce, hlavonožce, bentické živočichy s tvrdou schránkou aj.) nebo filtrovat plankton z vody. Po kořisti pátrají za pomoci několika smyslů, mezi nimiž hrají významnou roli čich, jemná mechanorecepce zprostředkovaná proudovým orgánem, jehož podstatnou součástí je postranní čára, a elektrorecepce zajišťovaná Lorenziniho ampulemi v kůži.
Nejstarší zkameněliny šupin, které lze považovat za šupiny paryb, jsou již ordovického stáří, kompletnější nálezy, které jsou k parybám řazeny s větší jistotou, pochází ze spodního siluru. Tyto rané paryby patří mezi tzv. trnoploutvé, což je skupina považovaná dříve za samostatnou třídu čelistnatců. Ukázalo se však, že je to parafyletická skupina, která z fylogenetického hlediska zahrnuje i nesporné paryby s chrupavčitou kostrou a dalšími pro ně typickými znaky. V mladších prvohorách byly jak chiméry (v širokém slova smyslu), tak příčnoústí velmi diverzifikovanými skupinami s mořskými i sladkovodními zástupci. Někteří byli hojní i v českých černouhelných pánvích (např. „pražralok“ Xenacanthus).
Popis



Paryby jsou pestrou skupinou čelistnatců, jejíž zástupci se vyskytují hlavně v mořích, vzácně i ve sladkých vodách. Mívají víceméně torpédovité nebo shora zploštělé tělo. Většinou dosahují středních až obřích rozměrů.[1] Za nejmenší z nich je považován parejnok narcina Hardwickova (Temera hardwickii), jehož samci mohou pohlavně dospívat již ve velikosti 8 cm;[2] největší parybou je žralok obrovský dosahující vzácně až 18 m.[1][3]
Chrupavčitá kostra
Jedním z nejvýznamnějších odvozených znaků definujících paryby je extrémní potlačení tvorby kostní tkáně. Prakticky celý jejich skelet je tak tvořen neosifikovanou chrupavkou, která je však především na povrchu zpevněná, mineralizovaná (kalcifikovaná). Právě chrupavčitá kostra byla motivací k odbornému pojmenování této skupiny: Chondrichthyes znamená doslova chrupavčité ryby (z řeckých slov χόνδρος, chrupavka a ἰχθύς, ryba);[4] v češtině se však pod označením chrupavčité ryby myslí spíše chrupavčití. Fosforečnan vápenatý je v povrchových částech chrupavek ukládán v podobě charakteristických šestibokých nízkých hranolů (prismat), které tak tvoří jakousi šestiúhelníkovou mozaiku (teselaci).[5][6][7] Výjimečně se kostní tkáň objevuje v neurálních obloucích obratlů.[5]
Lebka
Redukce kostní tkáně je u paryb spojena také s absencí krycích (dermálních) kostí,[5][7] které jinak u obratlovců patří k významným součástem lebky i jiných částí těla. Dermálního původu jsou např. kosti jako je horní a dolní čelist (tedy maxilla a mandibula = os dentale), kosti lebeční klenby (čelní, temenní kost, šupina spánkové kosti), nosní nebo skřelové kosti. Všechny tyto kosti u paryb chybí. Roli horní čelisti přebírá zejména chrupavčitá kost zvaná palatoquadratum (jež je u většiny obratlovců redukovaná, jeden z jejích pozůstatků se typicky jako tzv. kost čtvercová, os quadratum, podílí na stavbě čelistního kloubu, u savců je modifikována na kovadlinku), skelet dolní čelisti tvoří Meckelova chrupavka.[8] Na lebce nejsou patrné švy, její chrupavčité elementy kompletně srůstají. Vpředu bývá protažena ve zřetelný rypec (rostrum), pod nímž jsou uložena spodní ústa.[5][7]

Svalovina
Na alespoň částečně (u chimér zcela) zachovanou chordu a na páteř (u příčnoústých dobře vyvinutou a silně kalcifikovanou) a dále i na skelet ploutví, čelistí, žaberních oblouků aj. působí tělní svalovina zajišťující pohyb. Hlavní (tj. boční) sval je rozdělen na jednotlivé dobře zřetelné segmenty (myomery), které mají horní (epaxiální) část a dolní (hypaxiální) část, jež jsou odděleny horizontálně probíhajícím vazivovým septem.[9][10] Tak jako další obratlovci, i paryby mají jednak rychle reagující světlejší („bílá“, ve skutečnosti spíše růžová) svalová vlákna specializovaná na anaerobní rychlou glykolýzu, jednak efektivněji, ale pomaleji pracující a hůře unavitelná tmavší (červená) vlákna, která jsou lépe prokrvená a bohatá myoglobinem a mitochondriemi. Distribuce těchto typů svaloviny se liší podle převažujícího stylu pohybu jednotlivých druhů paryb.[10]


Ploutve
Síla svalů je přenášena mimo jiné na ploutve, hlavní orgány pohybu a jeho řízení. Paryby jsou vybaveny jednak nepárovými ploutvemi (v původní sestavě jsou to dvě hřbetní, jedna řitní a ocasní ploutev), jednak ploutvemi párovými (prsní a břišní ploutve). Zejména u rejnoků mohou být nepárové ploutve redukovány, naopak ploutve párové jsou u nich mohutné a mohou splývat do postranního lemu.[5] U některých žraloků může chybět jedna ze hřbetních ploutví (např. u šedounů) nebo řitní ploutev (např. u ostrounů).[7] Ploutve jsou vyztuženy elastickými proteinovými paprsky (keratotrichii).[5] Keratotrichia nejsou tvořena rohovinou, jak se v některých zdrojích uvádí. Mají povahu pevného vaziva, obsahují velké množství kolagenu produkovaného fibrocyty.[11] Prsní ploutve jsou ukotveny k pletenci tvořenému jedinou párovou chrupavkou (skapulokorakoid). Na skapulokorakoid navazují tři chrupavky zvané bazália, poté tzv. radiália a nakonec keratotrichia. Břišní ploutve jsou uchyceny k jediné příčné puboischiatické chrupavce.[12] Ocasní ploutev je zpravidla heterocerkní (s dominantním horním lalokem vyztuženým páteří), ale někdy je z vnějšku ocasní ploutev symetrická (a blíží se homocerkní ploutvi), u mnoha chimér a rejnoků je ocas bičovitý a tudíž vlastně difycerkní (i uvnitř symetrický). Vnitřní části břišních ploutví samců jsou přeměněny v kopulační orgány (pterygopody), oplození je totiž vnitřní.[5]

Kůže, šupiny
Kůže paryb je sama o sobě relativně tenká. Lze v ní identifikovat dvě základní vrstvy, povrchovou pokožku (epidermis) a škáru (dermis). Pokožka není tvořena zrohovatělými buňkami, ale živými buňkami chráněnými slizem produkovaným sekrečními buňkami. Další významný typ buněk v kůži jsou chromatofory (pigmentové buňky), které se významně podílejí na zbarvení.[9]
Tělo paryb je kryto zvláštním typem šupin, tzv. plakoidními šupinami. Jde o zahrocené šupiny se stavbou silně připomínající stavbu zubu: na povrchu je vrstvička skloviny (někdy zvaná vitrodentin) ektodermálního původu, pod ní se nachází zubovina (dentin), uvnitř je dutina s dobře prokrvenou dření. Šupina je ve škáře ukotvena destičkovitou bází, ve které se nachází pravá, i když bezbuněčná (acelulární) kostní tkáň. Plakoidní šupiny pronikají skrze pokožku a činí kůži paryb na omak drsnou (což je velmi zřetelné hlavně ve směru od ocasu k hlavě).[5][12] Rolí plakoidních šupin je kromě mechanické ochrany především snižovaní tření ve vodě. Typická plakoidní šupina má tři (nebo víc) vyvýšených ostrých kýlů oddělených hlubšími brázdami. Takové uspořádání potlačuje tvorbu mikrovírů ve vodě proudící podél šupin. Plakoidní šupiny u rychle plovoucích aktivních druhů jsou drobnější (kolem 300 μm), u pomalejších druhů mají až dvakrát větší rozměry.[10] Drsná kůže některých paryb, např. ostrounů, byla dříve využívána k hlazení dřeva podobně jako smirkový papír.[13] Několik skupin paryb plakoidní šupiny ztratilo (např. parejnoci či chiméry).[5][7]
Zuby a trávicí soustava
Stejnou stavbu jako plakoidní šupiny mají i zuby paryb. Zuby jsou trvale vyměňovány: nové zuby se zakládají v řadách podél čelistí uvnitř úst. Nejprve jsou skloněné vzad, postupně se posunují směrem vpřed, přičemž se zároveň na okraji čelistí napřimují a nahrazují opotřebované nebo vylomené zuby. Zuby mohou mít nejrůznější tvar: od trojúhelníkovitých ostrých, na okraji pilovitých zubů většiny žraloků přes zuby s několika hroty pro uchvácení kluzké kořisti, známé např. u žraloka límcového, až po ploché destičky řady rejnoků.[14] Někdy jsou zuby silně modifikované, např. u chimér jsou přeměněné v trvalé ploché desky s drticí funkcí.[5][12]
- Zuby a čelisti paryb
 Čelisti světlouna Bonnaterrova (Dalatias licha).
Čelisti světlouna Bonnaterrova (Dalatias licha). Dolní pohled na vnitřní plochu horní čelisti různozubce.
Dolní pohled na vnitřní plochu horní čelisti různozubce. Zadní pohled na čelisti a ploché zuby siby atlantské (Rhinoptera bonasus).
Zadní pohled na čelisti a ploché zuby siby atlantské (Rhinoptera bonasus).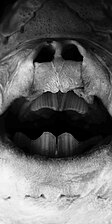 Pohled do otevřených úst chiméry.
Pohled do otevřených úst chiméry.


_1.jpg)

Trávicí trubice je relativně krátká a nepříliš objemná. Krátký jícen přechází v žaludek, na který navazuje tenké střevo s dobře vytvořenou spirální řasou (typhlosolis) zvětšující jeho vnitřní povrch.[5][9] U chimér je tato řasa vytvořena jen slabě.[15] Krátkým tlustým střevem (které je u paryb paradoxně užší nežli tenké střevo) ústí trávicí soustava do kloaky (u chimér má samostatný vývod). Trávicí trubice je doplněna pankreatem a objemnými, tukem vyplněnými játry.[9]

Dýchání a cévní soustava
U paryb na rozdíl od ryb nenacházíme plynový měchýř ani plíce. Roli hydrostatického orgánu nadnášejícího tělo mohou mít u pelagických druhů velká játra bohatá na tuky. Dýchání probíhá pouze pomocí žaber. Voda přes ně může u některých rychlých plavců proudit díky rychlému pohybu přes otevřená ústa, u řady zástupců však funguje nasávání vody přes ústa nebo modifikovanou první žaberní štěrbinu zvanou spirakulum (např. u rejnoků) a její následné protlačení přes žábry díky dýchacím pohybům čelistí a svalů ústní dutiny a žaber.[5]
Srdce je tvořeno čtyřmi dobře odlišitelnými za sebou (seriálně) uspořádanými oddíly: žilným splavem, jednou síní, jednou komorou a srdečním násadcem. Srdcem protéká odkysličená krev, která je pumpována do žaber, kde se obohatí o kyslík, a z nich pak teče dál do těla.[5]
Osmoregulace a urogenitální systém
Zajímavostí paryb je neobvykle vysoký obsah močoviny a trimethylaminoxidu, tedy sloučenin odvozených z toxického amoniaku, v krvi a dalších tělních tekutinách. Tyto látky zvyšují osmotický potenciál a tak přispívají k zadržování vody v těle – ta je totiž okolní slanou a hypertonickou mořskou vodou puzena ven z těla.[5][7] Močovina sice destabilizuje proteiny a je tudíž v takovýchto koncentracích toxická, trimethylaminoxid však funguje naopak jako stabilizátor proteinů a škodlivý efekt močoviny tak do značné míry eliminuje. I tak je pro paryby důležité udržovat vhodnou koncentraci iontů v těle, například se zbavovat nadbytečných sodných a chloridových iontů, které do jejich těla pronikají z mořské vody hlavně přes žábry, které jsou také důležitým osmoregulačním orgánem. Na osmoregulaci se u paryb kromě ledvin a žaber podílí také rektální žláza uložená nad zadní částí střeva, do něhož vylučuje přebytečnou sůl, která je pak vyloučena spolu s výkaly.[16] Ledviny, jimiž se paryby zbavují spíše přebytečné vody, jsou párové a podlouhlé, vpředu až vláknité, uložené podél hřbetu. Jejich tubuly mimo jiné vstřebávají zpět do těla výše zmiňovanou močovinu i trimetylaminoxid, aby byla zachována jejich vysoká koncentrace.[5][16] Ledvina je u samců zapojena i do procesu dozrávání a transportu spermií. Varlata jsou uložena vpředu na hřbetní straně tělní dutiny. Přední, vláknitá a stočená část ledviny zvaná též u paryb nadvarle (epidydimis; nejde o homolog savčího nadvarlete) zde přebírá spermie a transportuje je vzad do oblasti ledviny označované jako Leydigova žláza. Zralé spermie jsou pak uskladněny ve spermatických váčcích poblíž kloaky. U samic se ledviny na reprodukci nepodílejí. V přední části těla jsou vytvořeny vaječníky.[9] Ty jsou původně párové, ale u mnoha živorodých žraloků se zachovává jen jeden.[5] Zralá vajíčka jsou vejcovody, v nichž dochází k oplození, transportována do dělohy, která se během vývoje vajec nebo embryí zvětšuje.[9]

Rozmnožování
Jak bylo řečeno výše, u paryb dochází k vnitřnímu oplození za pomoci samčích pterygopodů. Paryby pak kladou buď poměrně velká vejce, nebo jsou vejcoživorodé až živorodé.[5] Paleontologická i fylogenetická data naznačují, že primitivní živorodost, kdy se sice rodí živá mláďata, ale i v těle matky jsou vyživována ze zásob ve žloutkovém váčku, je původním způsobem rozmnožování paryb (i u dnešních zástupců jde o častou variantu). Někdy je však embryo navíc vyživováno neoplozenými vejci poskytovanými mateřským organismem, výjimečně (jak je doloženo u žraloka písečného) se vyskytuje i nitroděložní kanibalismus, kdy nejsilnější embryo pozře své sourozence. U řady zástupců řádu žralouni se vytváří žloutková placenta, kdy se žloutkový váček postupně mění na orgán umožňující vstřebávat živiny přímo ze stěny dělohy.[7][17] Doba březosti u živorodých zástupců může být velmi dlouhá, u ostrounů trvá až 2 roky, což je nejdelší doložená březost mezi obratlovci.[5]


Smysly a nervová soustava
Zrak paryb zajišťovaný párem standardně stavěných komorových očí s kulovitou čočkou není příliš dobrý, ostrost vidění je relativně nízká a schopnost akomodace (přeostřování) je omezená.[5] V sítnici většiny paryb se nacházejí jak tyčinky, tak jediný typ čípků (jde o tzv. duplexní sítnici). Takové uspořádání prakticky neumožňuje rozlišování barev, ale přispívá patrně k velmi dobrému vnímání kontrastu. Jsou ale známy i druhy s dvěma či třemi typy čípků a schopností barevného vidění.[18]
Nozdry se u paryb zpravidla vyskytují jen jako jediný pár jamek na rostru, i když mohou být (většinou neúplně), přepaženy na dvě části (jednou voda proudí dovnitř, druhou ven). Čichový orgán uložený v kulovitých chrupavčitých čichových kapsulích v přední části lebky je tak omýván jednosměrným proudem vody. Vlastní čichový orgán (rozeta) je podobně jako u ryb tvořen mnoha lamelami (přepážkami), které mají povrch zvětšený ještě záhyby epitelu. Velikost, uspořádání i pozice čichového orgánu je však u paryb velmi variabilní a odrážejí styl života jednotlivých druhů. Za čichovým orgánem je uložen čichový bulbus, nápadný párový výběžek koncového mozku zpracovávající olfaktorické signály. U některých druhů je rozdělen na více částí.[18][19]
Obzvlášť žraloci mají pověst tvorů s vynikajícím čichem, ale ke kolujícím zkazkám o jejich skvělém čichu je třeba přistupovat s jistou dávkou opatrnosti; kriticky, experimentálně ověřených dat je pomálu. Např. se překvapivě ukazuje, že repertoár genů kódujících čichové receptory je u paryb (či přesněji u žraloků; situace u dalších skupin není dostatečně prozkoumaná) oproti jiným skupinám obratlovců značně redukovaný. Čich paryb vykazuje vysokou citlivost především k některým aminokyselinám a aminům, která je ale srovnatelná s citlivostí čichu paprskoploutvých ryb (jde o minimální zaznamenatelné koncentrace kolem 10−7–10−9 M, u některých žraloků i nižší). Vyšší citlivost by byla v zásadě zbytečná, protože zhruba v tomto rozmezí se pohybuje běžná koncentrace aminokyselin v moři tvořící jakési přirozené pachové pozadí. I tak je čich ve vodě důležitým smyslem. Vzhledem k tomu, že ve vodě je difuze méně účinná než ve vzduchu, udržují se molekuly pachů v daném vodním objemu déle a vodní proud, který takové pachy unáší, pak může vytvářet nápadnou, až několik kilometrů dlouhou pachovou stopu vedoucí ke kořisti. Právě parybami preferované aminosloučeniny se hojně uvolňují jednak z mrtvol zvířat, ale i z živé poraněné nebo právě požírané kořisti.[18][19] Nepříliš četné chuťové pohárky jsou lokalizovány pouze v ústech a hltanu (naproti tomu u ryb mohou být i na vouscích a jinde na povrchu těla).[5][18]
Vnitřní ucho s rovnovážnou i sluchovou funkcí a se třemi polokruhovitými kanálky je spojeno s vnějším prostředím tzv. endolymfatickým kanálkem (ductus endolymphaticus).[5][7] Na vnímání zvuku, tedy vibrací ve vodě, jakož i na vnímání proudění se podílí také proudový orgán. Jeho kanálky vytvářejí obzvlášť na hlavě komplikovaný systém, na těle pokračují jako postranní čára.[5] V proudovém orgánu jsou uloženy drobné mechanoreceptory (neuromasty), jimiž mohou paryby vnímat jemné změny tlaku a pohybu vody.[4] Zvuky vnímané parybami jsou spíše nízkofrekvenční (pro několik málo testovaných druhů žraloků v rozsahu asi 20–300 Hz). V jednom z experimentů přilákaly žraloka zvuky napodobující hluk vydávaný mrskající se rybou na vzdálenost až 800 m.[20] Kromě vibrací se ve vodě spolu se zvukem produkovaným kořistí vyskytují v blízkém okolí zdroje skoro vždy i proudy, případně víry vyvolané pohybem zdroje. Toto proudění je zaznamenáváno proudovým orgánem podél těla paryby.[18]
Zajímavým smyslem paryb je elektrorecepce zprostředkovaná drobnými četnými Lorenziniho ampulemi rozesetými po hlavě. Díky nim mohou vnímat i velmi slabá elektrická napětí generovaná např. činností svalů kořisti.[5][21] Kromě toho využívají např. rejnoci elektrorecepci i při vyhledávání partnerů. Další druhy paryb (nebo i embrya ve vejci) díky ní rozpoznají blížícího se predátora a mohou dočasně strnout, aby neupoutaly pozornost. U trnuch byla prokázána schopnost vnímat magnetické pole. Nakolik se na těchto vjemech podílejí Lorenziniho ampule nebo jiné magnetoreceptory, není jasné.[18]
V mozku je dobře vyvinut koncový mozek (konkrétně jeho čichové laloky) a mozeček, který řídí koordinaci pohybů a pomáhá i s udržováním správné polohy. Podněty z očí, vnitřního ucha i z proudového orgánu jsou zpracovávány ve středním mozku.[5]
Endokrinní soustava
Co se žláz s vnitřní sekrecí týče, je u paryb vyvinuta štítná žláza, hypofýza, nadledviny (ve formě dvou samostatných žláz odpovídajících dřeni a kůře nadledvin) nebo pankreas s endokrinní i exokrinní funkcí.[5]
Systém a diverzita
Paryby jsou dnes zastoupeny dvěma podtřídami, které se nazývají příčnoústí (Elasmobranchii nebo též Neoselachii) a chiméry čili celohlaví (Holocephali).[5][22]
Chiméry mají velkou hlavu a zvětšené prsní ploutve, které jsou hlavním orgánem pohybu. Před první hřbetní ploutví se nachází trn, jejich ocas je většinou (až na chimérovkovité) štíhlý a někdy až bičovitě protažený, tedy de facto difycerkní. Mají vytvořenou nepravou kožní skřeli, která překrývá pouhé čtyři funkční žaberní štěrbiny. Spirakulum je uzavřeno. Jejich lebka je kompaktní, tzv. holostylní, palatoquadratum je přirostlé k neurokraniu (odtud označení „celohlaví“ – Holocephali). Zuby srůstají do plochých desek a nejsou vyměňovány. Obratle jsou bez těl a chorda je tudíž kompletně zachovalá. Nemají vytvořenou kloaku, trávicí a urogenitální soustava mají samostatné vývody.[5] Chiméry jsou zastoupeny asi 50 recentními druhy.[4]
Příčnoústí jsou mnohem pestřejší a početnější skupina než chiméry. Lze je rozdělit na žraloky (Selachii), mezi něž ale patří také např. polorejnoci a pilonosové, a rejnoky (Batoidei) včetně pilounů, parejnoků a dalších.[23] Žraloků je známo přes 500 druhů, rejnoků necelých 700.[4] Příčnoústí mají rozmanitou stavbu těla: to je buď torpédovité, nebo shora zploštělé a rozšířené. Palatoquadratum je k neurokraniu připojeno volněji než u chimér: buď prostřednictvím vazů (tzv. amfistylní lebka) nebo přes kost, u paryb samozřejmě chrupavčitou, zvanou hyomandibulare (hyostylní lebka).[5] Žaberní štěrbiny (nejčastěji v počtu pěti, výjimečně 6–7 párů) u nich vyúsťují samostatně a spirakulum bývá zpravidla zachováno (ač ne vždy). Obzvlášť u rejnoků je dobře vyvinuto a slouží k nasávání vody při dýchání.[24] Do chordy zasahují kalcifikovaná těla obratlů, takže je zaškrcovaná resp. přerušovaná.[5]
Rejnoci jsou i v relativně recentní české literatuře většinou pojímáni jako jediný řád (Rajiformes),[1] moderněji jsou ale děleni na minimálně 4 řády: rejnoci (Rajiformes), trnuchy (Mylobatiformes), pilouni (Rhinopristiformes) a parejnoci (Torpediniformes).[7][23] Dvě základní evoluční větve žraloků jsou Squalomorphi a Galeomorphi. Squalomorphi jsou anatomicky poněkud primitivnější, mají menší mozek než Galeomorphi a jsou více vázáni na hlubší vody, kde asi vznikli. Galeomorphi jsou hojnější v tropických pobřežních vodách.[25] Je rozlišováno pět řádů squalomorfních a čtyři řády galeomorfních žraloků.[23] Blíže viz tabulka níže.
V následující tabulce je shrnut přehled systému recentních paryb. Je založena hlavně na publikaci Nelsona et al. (2016)[7] a doplněna na základě dalších zdrojů.[23][24][26]
| Třída paryby (Chondrichthyes) | |||
| Podtřída celohlaví (Holocephali) | |||
| Řád | Hlavní znaky | Čeledi | Ukázka |
|---|---|---|---|
| Chiméry (Chimaeriformes) | Mohutné prsní ploutve. První hřbetní ploutev s trnem, který lze vztyčit, druhá hřbetní ploutev nízká a dlouhá. Pro přívod vody k dýchání slouží i nozdry. | Chimérovkovití (Callorhynchidae), pachimérovití (Rhinochimaeridae), chimérovití (Chimaeridae) | 
pachiméra atlantská (Rhinochimaera atlantica) |
| Podtřída příčnoústí (Elasmobranchii) | |||
| Nadřád žraloci (Selachii), skupina Squalomorphi | |||
| Řád | Hlavní znaky | Čeledi | Ukázka |
| Šedouni (Hexanchiformes) |
Zvýšený počet žaberních štěrbin (6–7). Jediná hřbetní ploutev umístěná dosti daleko vzadu. Drobné spirakulum leží daleko za okem. |
Štíhlounovití (Chlamydoselachidae), šedounovití (Hexanchidae) | 
žralok Nakamuraiho (Hexanchus nakamurai) |
| Ostrouni (Squaliformes) |
Chybí řitní ploutev. Dvě hřbetní ploutve, s ostny či bez ostnů. Spirakulum vytvořeno. |
Bezkýlovcovití (Centrophoridae), Etmopteridae, Somniosidae, Oxynotidae, světlounovití (Dalatiidae), ostrounovití (Squalidae) | 
ostroun obecný (Squalus acanthias) |
| Echinorhiniformes |
Poslední z pěti žaberních štěrbin je větší než ostatní. Malé spirakulum dalece za očima. Dvě malé hřbetní ploutve bez trnů, řitní ploutev chybí. |
Drsnotělcovití (Echinorhinidae) | 
žralok Cookeův (Echinorhinus cookei) |
| Polorejnoci (Squatiniformes) |
Bentické paryby. Zploštělé tělo, mohutné párové ploutve. Řitní ploutev chybí. Oči orientované směrem nahoru. Dobře vyvinuté spirakulum. |
Polorejnokovití (Squatinidae) | 
polorejnok kalifornský (Squatina californica) |
| Pilonosi (Pristiophoriformes) |
Protažené tělo, rypec dlouhý, plochý, po stranách ozubený nestejně velikými zuby. Vytvořen pár dlouhých vousků, chybí řitní ploutev. Spirakulum velké. |
Pilonosovití (Pristiophoriformes) | 
pilonos japonský (Pristiophorus japonicus) |
| Nadřád žraloci (Selachii), skupina Galeomorphi | |||
| Řád | Hlavní znaky | Čeledi | Ukázka |
| Různozubci (Heterodontiformes) | Dvě hřbetní ploutve s trny. Vysoká hlava, oči vysoko, spirakulum drobné, nozdry propojeny s ústy hlubokou rýhou. Zadní zuby ploché. Šroubovitý vzor na povrchu vajec. | Různozubcovití (Heterodontidae) | .JPG)
různozubec Francisův (H. francisi) |
| Malotlamci (Orectolobiformes) | Standardní sestava ploutví (dvě hřbetní bez trnů, řitní přítomna), spirakulum vytvořeno, umístěno nedaleko za okem. Od nozder k ústům vede nápadná rýha, časté vousky. | Kobercovcovití (Parascyllidae), slepounovití (Brachaeluridae), wobegongovití (Orectolobidae), žralůčkovití (Hemiscylliidae), vouskatcovití (Ginglymostomatidae), pruhovcovití (Stegostomatidae), veležralokovití (Rhincodontidae) | 
žralůček šedý (Chiloscyllium griseum) |
| Obrouni (Lamniformes) | Standardní sestava ploutví, bez vousků, ústa zasahují až pod oko. Malé spirakulum zpravidla vyvinuto. Střevní spirální řasa ve formě řady kroužků. Časté požírání vajec jako forma výživy embryí. | Hlavorohovití (Mitsukurinidae), písečníkovití (Odontaspididae), krokodýlovcovití (Pseudocarchariidae), liškounovití (Alopiidae), velkotlamcovití (Megachasmidae), obrounovití (Cetorhinidae), lamnovití (Lamnidae) | 
žralok písečný (Carcharias taurus) |
| Žralouni (Carcharhiniformes) | Standardní sestava ploutví. Chybí žaberní tyčinky. Ústa protažena vzad až pod oko. Pohyblivé dolní oční víčko tvoří mžurku chránící oko. Spirakulum zpravidla chybí. | Máčkovití (Scyliorhinidae), pruhounovití (Proscylliidae), mnohozubcovití (Pseudotriakidae), vousatcovití (Leptochariidae), hladkounovití (Triakidae), velkookatcovití (Hemigaleidae), modrounovití (Carcharhinidae), kladivounovití (Sphyrnidae), Pentanchidae | 
žralok černoploutvý (Carcharhinus melanopterus) |
| Nadřád rejnoci (Batoidei) | |||
| Řád | Hlavní znaky | Čeledi | Ukázka |
| Parejnoci (Torpediniformes) | Z žaberní svaloviny vytvořeny orgány generující elektrické napětí. Kůže měkká a volná. Oči drobné. Dobře vyvinutá ocasní ploutev. | Parejnokovití (Torpedinidae), narcinovití (Narcinidae), Platyrhinidae? | 
parejnok elektrický (Torpedo marmorata) |
| Rejnoci (Rajiformes) | Ocasní ploutev někdy zakrňuje, ocas je štíhlý. Mnohdy mají ostny na povrchu (zvětšené plakoidní šupiny), často uspořádané podél hřbetu. Silně vyvinuté, vzadu zploštělé pterygopody u samců. | Rejnokovití (Rajidae) | 
rejnok paprsčitý (Amblyraja radiata) |
| Pilouni (Pristiformes nebo Rhinopristiformes) | Tělo jen částečně zploštělé, není tak široké jako u ostatních rejnoků, ocas není příliš štíhlý a tak zřetelně oddělený od těla. Více či méně protažený rypec. | Pilohřbetovití (Rhinobatidae), kytarovcovití (Rhinidae), Rhynchobatidae, Zanobatidae, pilounovití (Pristidae) | ,_auch_Gewöhnlicher_Geigenrochen_oder_Gewöhnlicher_Gitarrenfisch.DSCF9461WI.jpg)
pilohřbet obecný (Rhinobatos rhinobatos) |
| Trnuchy (Myliobatiformes) | Na ocase vytvořen jeden nebo více pilovitých trnů asociovaných často s jedovou žlázou (u některých zástupců sekundárně chybí). Chybějí hrudní žebra. Typicky mají větší mozek. | Hlubinářovití (Plesiobatidae), tlustoocaskovití (Urolophidae), šestižábrovcovití (Hexatrygonidae), trnuchovití (Dasyatidae), Potamotrygonidae, křídlounovití (Gymnuridae), Urotrygonidae, mantovití (Myliobatidae) | 
trnucha tečkovaná (Dasyatis guttata) |
Ekologie

Paryby jsou především mořští živočichové, několik desítek druhů žije ve sladkých nebo brakických vodách, případně ze slaných vod vplouvá do řek. V tropech je větší diverzita paryb nežli v chladnějších oblastech. Mezi parybami lze nalézt jak aktivní, často vrcholové predátory („typické“ žraloky lovící ryby, hlavonožce nebo mořské savce a ptáky), tak i planktonofágní, někdy obrovské druhy (např. žralok obrovský nebo ž. veliký, ale také manty) nebo bentické lovce číhající skrytě na kořist (polorejnoci aj.) či sbírající bezobratlé ze dna, často se zuby specializovanými na drcení jejich schránek (např. někteří rejnoci nebo chiméry).[5]
Paryby se rozmnožují pomalu, rodí malý počet mláďat nebo snášejí menší množství velkých vajec (jsou to K-stratégové). Velké druhy pohlavně dospívají ve věku kolem 15–30 let. I z těchto důvodů jsou paryby zranitelné lovem i dalšími lidskými činnostmi a lze mezi nimi nalézt i vážně ohrožené druhy.[5]
Paleontologie


V recentní době (ve významnějším rozsahu asi od 10. let 21. století) došlo k významnému posunu názorů na nejstarší historii paryb. Vymřelá skupina trnoploutvých (Acanthodii), která byla považována za samostatnou skupinu archaických čelistnatců, začala být na základě výsledků fylogenetických analýz vnímána jako parafyletický vývojový stupeň (grad) vedoucí právě k typickým parybám. Trnoploutví jsou tedy považováni za zástupce tzv. kmenové linie paryb a jako primitivní paryby jsou také interpretováni. Na některých lépe zachovaných fosiliích jsou patrné přechody mezi „typickými“ trnoploutvými a ranými „pravými“ parybami; žádná ostrá hranice mezi oběma skupinami neexistuje.[27][28][29] Dobře prozkoumanou přechodnou formou mezi klasickými trnoploutvými a typickými parybami je devonský Doliodus s mozaikou znaků přisuzovaných dříve buď trnoploutvým (charakter šupin, trny před prsními ploutvemi), nebo parybám (kalcifikovaná prizmatická chrupavka, stavba pletence prsních ploutví).[30][28] Mezi nejstarší dobře doložené zástupce trnoploutvých/paryb patří Fanjingshania ze spodnosilurských čínských vrstev (asi 439 miliónů let starých)[31] nebo jen o málo mladší Shenacanthus (436 miliónů let);[32] oba rody jsou popsané velmi recentně (r. 2022). Další fragmentární pozůstatky (šupiny a ploutevní trny řazené do taxonů Mongolepidida resp. Sinacanthida), které lze s větší či menší mírou nejistoty přiřadit trnoploutvým parybám, jsou známy dokonce již z ordoviku.[33]

Významný devonský rod byl Cladoselache (přestože je běžně označován – a dříve i považován – za prvohorního žraloka, ve skutečnosti nepatří do nadřádu žraloci ani mezi příčnoústé, fylogeneticky je zřejmě bližší chimérám).[27][34] Byla to paryba dlouhá až 2 m s prakticky symetrickou ocasní ploutví (ačkoli chorda a páteř zabočovaly do horního laloku); pravděpodobně šlo o rychlého plavce.[29] Přinejmenším před přední hřbetní ploutví byl silný trn.[29][35] Příbuzný rod Maghriboselache možná vykazoval i nápadný sexuální dimorfismus: přední hřbetní ploutev s výrazným trnem asi byla přítomná pouze u samců, jimž mohla sloužit při páření.[36] Podobnou roli snad hrály i silně modifikované trny první hřbetní ploutve u samců některých paryb ze skupiny Symmoriida považované někdy za příbuzné příčnoústých,[37] v recentnějších studiích však spíše celohlavých.[34][36] Např. samci rodu Falcatus měli vpředu na hřbetě vytvořen zvláštní, vpřed nad hlavu ohnutý plochý útvar, samci rodů Stethacanthus nebo Akmonistion nesli na hřbetě bizarní útvar, který se směrem vzhůru se rozšiřoval, nahoře byl plochý a pokrytý zoubky.[37]
S mnohem větší jistotou jsou do velmi pestrého příbuzenského okruhu celohlavých řazeni např. zástupci fosilních řádů Iniopterygiformes s mohutnými prsními ploutvemi, Chondrenchelyformes s dlouhým úhořovitým tělem, Eugeneodontiformes s velmi zvláštní zubní spirálou nebo Petalodontiformes s bachratým či zploštělým tělem a zuby specializovanými na drcení. Přímo mezi celohlavé je jako raný zástupce řazen např. sladkovodní permo-karbonský rod Helodus.[37]
- Fosilní zástupci evoluční linie směřující k chimérám
 Sibyrhynchus (Iniopterygiiformes)
Sibyrhynchus (Iniopterygiiformes) Chondrenchelys (Chondrenchelyformes)
Chondrenchelys (Chondrenchelyformes) Helicoprion (Eugeneodontiformes): zubní spirála
Helicoprion (Eugeneodontiformes): zubní spirála Strigilodus (Petalodontiformes)
Strigilodus (Petalodontiformes)




K archaickým zástupcům příčnoústých patří např. sladkovodní skupina Xenacanthiformes známá od devonu do triasu. Význačný rod Xenacanthus známý i z českého permu měl protažené tělo, dlouhou hřbetní ploutev, dále postupně se zužující, v podstatě difycerkní (vně i uvnitř symetrickou) ocasní ploutev a dlouhý trn za hlavou.[12][37] Patrně se dobře pohyboval jezery zarostlými bujnou vegetací. Další starobylé řády jsou Ctenacanthiformes nebo Hybodontiformes. Hybodontiformes byli pestrou skupinou s různými typy zubů poukazujícími na různé potravní specializace, původně byli mořští, ale v někteří přešli ke sladkovodnímu způsobu života. V juře byli dominantní skupinou, v křídě koexistovali s bouřlivě se rozvíjejícími moderními příčnoústými (s jejich korunovou skupinou, tedy s Neoselachii).[37]
Neoselachii byli díky pevnější (kalcifikované) páteři schopni využívat větší sílu svaloviny a tak dosahovat při pohybu vyšší rychlosti než jejich fosilní příbuzní, mohli šířeji otevírat tlamu díky volnějšímu palatoquadratu a tak efektivněji lovit (mimo jiné i díky podtlaku a sání, ke kterému dojde v důsledku náhlého otevření úst), vyvinuly se u nich jemně pilovité zuby schopné porcovat kořist. Celkově šlo tedy o zdatné predátory. Mají i větší mozek než archaičtější skupiny, některé smysly jsou u nich zřejmě také vyvinutější (mají větší čichové orgány než vymřelé skupiny paryb).[38] První z nich (šedouni, různozubci) se objevují koncem triasu.[39] Asi nejslavnější fosilní zástupce moderních příčnoústých je obrovský, podle různých odhadů 15–16,[40][41] možná až 20 m[42] dlouhý neogenní superpredátor, megalodon.[43]
- Vyhynulí zástupci příčnoústých
 Orthacanthus, velký (až třímetrový) zástupce řádu Xenacanthiformes
Orthacanthus, velký (až třímetrový) zástupce řádu Xenacanthiformes
 Zub megalodona
Zub megalodona Fosilní rejnok Heliobatis radians
Fosilní rejnok Heliobatis radians




Odkazy
Wikiwand - on
Seamless Wikipedia browsing. On steroids.