オパビニア
カンブリア紀の節足動物 ウィキペディアから

オパビニア(Opabinia[4])は、約5億年前のカンブリア紀に生息したオパビニア類[5]の古生物の一属。体の先頭に突出したハサミ状の吻と5つの眼、両筋にたくさんの鰭と鰓をもつ[6][7][8][9][10][5]。カナダのバージェス頁岩で見つかったオパビニア・レガリス(Opabinia regalis[4])という1種のみ知られている[6][7][11][5]。
| オパビニア | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 オパビニアの復元図 | |||||||||||||||||||||
| 保全状況評価 | |||||||||||||||||||||
| 絶滅(化石) | |||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||
| 古生代カンブリア紀ウリューアン期 (約5億1,000万 - 5億500万年前)[1] | |||||||||||||||||||||
| 分類 | |||||||||||||||||||||
| |||||||||||||||||||||
| 学名 | |||||||||||||||||||||
| Opabinia Walcott, 1912 [4] | |||||||||||||||||||||
| タイプ種 | |||||||||||||||||||||
| オパビニア・レガリス Opabinia regalis Walcott, 1912 [4] |
かつてはその独特な姿により、バージェス動物群の中で現存する動物の分類群には収まりきらない「プロブレマティカ」(未詳化石)と疑問視され、「奇妙奇天烈動物」(weird wonders)の代表例として語られてきた有名な一生物である[12][10]。後に研究が進み、アノマロカリスなどのラディオドンタ類、ケリグマケラ、パンブデルリオンと共に、基盤的な節足動物として広く認められるようになった[3][8][13][9][10][2][14][15][16][17][5]。
名称
学名「Opabinia」は、発見地(カナダブリティッシュコロンビア州、バージェス頁岩、ウォルコットとレイモンド採石場)近くのオハラ湖(Lake O'Hara)の南東部にある、ヨーホー国立公園(Yoho National Park)のハンガビー山(Mount Hungabee)とビッドル山(Mount Biddle)の間に位置するオパビン峠(Opabin Pass)に由来する(「Opabin」は現地の言葉で「岩」を意味する[18])[4]。模式種(タイプ種)の種小名「regalis」はラテン語で「王の」を意味する[11]。
化石と発見

オパビニアの化石標本は希少で[11]、カナダブリティッシュコロンビア州のバージェス山にある堆積累層、バージェス頁岩(Burgess Shale、カンブリア紀ウリューアン期、約5億1,000万 - 5億500万年前[1])のウォルコットとレイモンド採石場(Walcott and Raymond Quarries)のみから唯一の種オパビニア・レガリス(Opabinia regalis)が発見される[11]。本属はアメリカ古生物学者チャールズ・ウォルコット(Charles Doolittle Walcott)によって最初に発見され、1912年に記載された[4]。しかし、その独特な姿が解明され、多くの注目を集めるようになったのは、イギリスの古生物学者ハリー・ウィッティントン(Harry B. Whittington)に再記載が行われた1970年代以降である(後述)[6][10]。選定基準標本(レクトタイプ)USNM 57683 はアメリカの国立自然史博物館(Smithsonian National Museum of Natural History)に所蔵される[11]。
シベリアで見つかり、Miroshnikov & Krawzov 1960 に本属の種(Opabinia norilica)として記載された化石標本もあった[19]が、これは後の研究に本属由来のものとして認められない[6]。
形態
要約
視点
 オパビニア・レガリス(Opabinia regalis)の全身復元図
オパビニア・レガリス(Opabinia regalis)の全身復元図 同種のサイズ測定図
同種のサイズ測定図


体長は吻を除いて4 - 7cm程度[11]。1本の吻に融合したハサミ型の前部付属肢[7][20]・眼柄に突出した5つの眼・十数対の鰭(ひれ)と鰓(えら)を有する長い胴部を掛け合わせた独特な姿をもつ[6][8]。眼・鰓・体節・消化管などの部分は節足動物的だが、表皮(クチクラ)は柔軟で、硬質の外骨格ではない[8][13][2]。基本体制は同じオパビニア類のユタウロラに共通し、オパビニア類に近縁とされるラディオドンタ類(アノマロカリスなど)、ケリグマケラとパンブデルリオンにも似ているが、前部付属肢・口・鰭など一部の構造はやや異なる[7][8][21][20][13][22][2]。
頭部
頭部は胴部に対して明らかに小さく、特徴的な眼と吻はそれぞれ前上方と前下方にある(後述)[6][7][8]。腹面はやや後方に膨らんで、後ろに向けて開いた口をもつ[6][7][8]。口の周辺は放射状の構造体に囲まれ、ラディオドンタ類の口器(oral cone)を思わせるが、そのような硬質の歯の有無ははっきりしない[20]。
眼
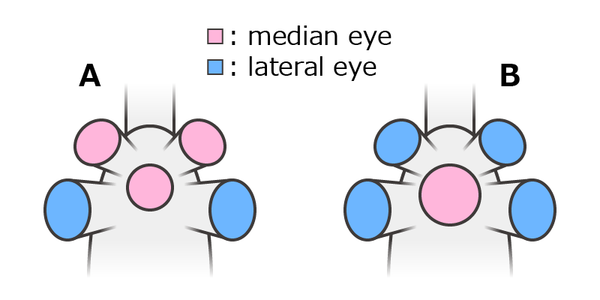 オパビニアの中眼(ピンク)と側眼(青)に対する2つの解釈
オパビニアの中眼(ピンク)と側眼(青)に対する2つの解釈
A: 小型中眼3つ側眼1対、B: 大型中眼1つ側眼2対

頭部の前上方にある大小5つの眼は前方1対・両後方1対・中央1つのパターンで配置される。前後2対の眼はどれも丈夫な眼柄に突出した複眼で、両後方の1対は前方の1対より大きい[6][8][23]。
似たような眼をもつキリンシアと同様、これらの眼は節足動物において一般的な2種類の眼、すなわち中央にある小さな中眼(median eye)と左右にある大きな側眼(lateral eye)に相同とされる[23]。しかし前方1対の眼の相同性と中央1つの眼の形態は、文献により解釈がやや異なる。通説では全ての眼が眼柄をもち、両後方の1対のみ側眼で、残りの3つはどれもやや小さな中眼だとされる[23]。一方、Moysiuk & Caron 2022 では中央の1つのみ中眼で、眼柄をもたず、従来の復元より大きかったと解釈される[24]。
吻(前部付属肢)


頭部の前下方にある1本の吻(proboscis)は体長の3分の1ほど長く、表皮は無数の環形の筋(annulation)に細分される[6][7]。可動域は広く、正面に伸びた姿勢と体の腹面に折り畳んだ姿勢で化石に保存されることが多い[6]。吻の先端にある左右1対の短い前部付属肢(frontal appendages)は、それぞれの先端に少なくとも5本の細い爪があり、横開きのハサミに似た捕獲器をなしている[7][8][20]。
この吻は一部の節足動物の関節肢(例えば昆虫の下唇)のように、元々1対だった付属肢が、基部を左右融合することにより特化した部分であり[7][25][26][27]、前部付属肢の機動性を高めるように進化したものだと考えられる[20]。これは一見してラディオドンタ類、ケリグマケラとパンブデルリオンの左右独立した前部付属肢とは大きく異なるが、棘をもつ、眼と口より前に特化した付属肢であることが共通している[7][25][3][20][2]。
Whittington 1975 をはじめとして[6]、この吻の前部付属肢は誤って異様な縦開きに復元されることが多い[12][11]。しかしこの部分は横開きであることが背腹(USNM 57684, USNM 205258, USNM 155599, YPM 5809)と側面(USNM 57683)に保存された複数の化石標本で明確に示されている(背腹だと対になる、側面だと重なる)のに加えて、左右対称の形・発生学的観点・近縁(ラディオドンタ類、ケリグマケラとパンブデルリオン)の前部付属肢との比較も、横開きが合理的であることを示している[7][25][8][20][27]。
胴部


黄色:消化管
オレンジ色:消化腺
暗青色:鰓
水色:鰭
緑色:葉足(不確実)
胴部は円筒状に長く、わずかに湾曲できる15節の胴節と特化した尾部が含まれる[6][7][8]。硬質の外骨格(背板と腹板)はなかった[2]ものの、各胴節の境目は背面で顕著に見られる[6][7][8][13]。各胴節の付属肢由来と思われる構造としては、発達した鰭、櫛状の構造体(鰓)、および有無が議論的な脚が知られている[9]。これらの構造は、再記載ごとに解釈がやや異なる(詳細はオパビニア#鰭・鰓の構造と脚の有無を参照)[6][7][8][13][9][10]。
鰭と鰓
15対の鰭(flaps, lobes)は丸みを帯びた長方形で、各胴節の左右から生えている[6][8][22]。化石では様々な角度に保存されても顕著に見られることから、これらの鰭は(ラディオドンタ類、パンブデルリオンとケリグマケラのような)水平ではなく、斜め下に向けて張り出したと考えられる[6][8]。隣接した前後の鰭は重なった部分があり、後ろの鰭の前縁は直前の鰭の後縁に覆われる[6][8][22]。それぞれの鰭の表面は、鰓(えら)と思われる一面の櫛状の構造体(setal blades, setal structures)がある[8][9]。これはたくさんの細長い葉状の附属体(lanceolate blades, gill blades)によって構成され、鰭の前縁近くある1本の筋に沿って連結していた[8][9]。
これ似た鰭と鰓は、ユタウロラ、ラディオドンタ類、ケリグマケラとパンブデルリオンにも見られる[7][25][8][28][21][20][13][9][22][5]。しかし、ユタウロラの鰓は鰭の一部と胴節を同時に覆うこと、ラディオドンタ類の鰭の畳み方は逆で鰓は鰭を覆わない(胴節のみを覆う)こと、およびパンブデルリオンとケリグマケラの鰓は(細長い葉状よりも)細い繊毛状であることが、オパビニアのものとは明確に異なる[8][22][5]。これらの古生物、特にオパビニア、ユタウロラとラディオドンタ類の鰓は、他の節足動物の外葉(exite)に相同だと考えられる[8][13][9][22][2][29][5]。
脚(葉足)
Budd 1996 をはじめとして、鰭に覆われた各胴節の腹面はパンブデルリオンとケリグマケラのように、葉足(lobopod, 葉足動物などに見られる柔軟な付属肢)という脚をもつ可能性が示された[8]。しかしこれは Zhang & Briggs 2007 をはじめとして疑問視され[13]、その有無は2010年代現在では未だに確実でない[9][10][22]。もし脚が存在すれば、それは円錐状の短い付属肢であり、表皮は環形の筋に分かれ、先端に爪はない[9]。この脚は、ラディオドンタ類の腹側の鰭、他の節足動物の内肢(endopod)、およびそれ以外の汎節足動物(葉足動物・有爪動物・緩歩動物)の葉足に相同だと考えられる[8][22]。
尾部
最終胴節に続く尾部は胴節の3倍ほど長く、左右から斜め上に特化した3対の尾鰭(tail fan blade)があり、発達した尾扇(tail fan)を構成する[6][7][8]。畳み方は胴節の鰭とは逆で、尾鰭は前の鰭の後縁が直後の鰭の前縁に覆われる[6][7][8]。体節の境目は見当たらない[6]が、尾鰭の数から、この尾部は少なくとも3節の体節を含んだと考えられる[7]。最終の尾鰭の間に当たる末端は1対の目立たない尾毛(caudal rami)で、その縁が鋸歯状になっている[5]。
このような尾部は同じオパビニア類であるユタウロラ[5]の他にラディオドンタ類にも見られ、その中でもアノマロカリスのものがよく似ている[25][8][3][30]。
内部構造

消化管が明瞭に発見されている[6][31]。口は頭部の腹側で後ろ向きに開くため、消化管の前端(咽頭)はUターンして折り返している[6][2][14]。第2胴節から第13胴節にかけて、中腸の左右に11対の丸い消化腺(digestive gland、中腸線 midgut gland)が並んでいる[9][31]。このような消化系は節足動物的であり、複数対の消化腺は早期の節足動物(シベリオン類[32][33]、ケリグマケラ[34]、パンブデルリオン[28]、ラディオドンタ類[35][36][37]、キリンシア[23]、イソキシス[38]、フーシェンフイア類[39]、メガケイラ類[40]、三葉虫[41]など)を通じて一般に見られ[31][2][14][16]、折り返した咽頭(=後ろ向きの口)は真正の節足動物に共通の特徴でもある[2][14]。
消化管以外では、体腔(body captivity)であった可能性が高い、各胴節から三角形に枝分かれた痕跡が知られている[8][9]。
生態


オパビニアは遊泳底生性(nektobenthic, 底生性に近い遊泳性)の捕食者であったと考えられる[6]。バージェス頁岩に保存された化石は堆積物の埋蔵に由来するため、オパビニアは主に海底に生息したと推測される(でなければそのように埋蔵される可能性が低い)[6]。鰓をもつ各胴節の鰭を上下に動かすことにより、海底の表面で緩く遊泳しながら呼吸もできたと考えられる[6]。胴部の可動域は低いため、体のうねりで遊泳することはできなかったと考えられる[6]。なお、もし各胴節の鰭を順序よく波打つことができれば、オパビニアは前述の予想以上に高い遊泳能力をもつ同時に、尾部の尾扇をステアリングに用いていたと考えられる[6]。
多数の複眼は発達で様々な方向に向くため、オパビニアは良好な視力と広い視野を有し、上方・側方の光や動きの変化を察するに適したと考えられる[6]。能動的な吻は筋肉質であったと推測され、堆積物から餌を見つけ出し、先端の前部付属肢でそれを捕獲しては、そのまま後ろ向きの口まで運ぶことができたと考えられる[6]。口に発達した硬質の口器はなかったため、餌は柔軟であったと推測される[6]。特化した消化腺をもつことにより、餌を効率よく消化することが出来たと考えられる[31]。
研究史と分類史
要約
視点
オパビニアは、カンブリア紀の古生物の中でも形態学上の復元と分類学上の位置付けに多くの議論を繰り広げられた一生物である[10]。20世紀初期では甲殻類[4]、20世紀後期では議論的な未詳化石(プロブレマティカ)[12]、1990年代以降ではオパビニア類の一員としてアノマロカリスなどのラディオドンタ類と共に、基盤的な節足動物として広く認めされるようになった[8][21][13][9][10][2][14][15][16][17]。
甲殻類(1910~1970年代)


バージェス頁岩(バージェス動物群)から最初に見つかった頃ではその特異性が理解されておらず、オパビニアはほぼ疑いもなく甲殻類の節足動物だと考えられた[4]。原記載である、アメリカの古生物学者チャールズ・ウォルコット(Charles Doolittle Walcott)の1912年の記載(Walcott 1912)では、オパビニアは現存するThamnocephalidae科の無甲類(鰓脚類に分類される、ホウネンエビとアルテミアを含んだ甲殻類の分類群)と比較され、それに似た原始的な無甲類と見なされた。また、この類の無甲類のオスは先頭に枝分かれた吻があるため、それを思わせる吻(前部付属肢)が見られるオパビニアの化石標本は全てオスだと解釈された[4]。なお、オパビニアの化石標本から無甲類にあるはずの頭部構造(2対の触角・大顎・小顎)が見当たらず、鰭(当時では「胸肢」とされた)にある櫛状構造の解釈に難点があることも明記された[4]。


この見解は動物学者ジョージ・イヴリン・ハッチンソン(George Evelyn Hutchinson)にも踏襲され、Hutchinson 1930 では、ホウネンエビのように背泳ぎの甲殻類として解釈された、史上初[10]のオパビニアの復元図が載せされた[42]。Simonetta 1970 では、オパビニアの化石標本に見当たらないものの、当時ではオパビニアの近縁と誤解釈されたレアンコイリアとヨホイア(後述)の特徴に基づいて、背板・触角・関節に分かれた脚などという節足動物的な特徴をオパビニアの復元図に多く追加された[43]。
未詳化石(1970~1980年代)

1970年代、イギリスの古生物学者ハリー・ウィッティントン(Harry B. Whittington)の再記載(Whittington 1975)により、オパビニアは甲殻類どころか、一見して節足動物にすら分類しにくいほど独特な特徴をもつことが分かった[6]。これは1972年にバージェス動物群の学会発表があった際、ウィッティントンのこの復元図が映し出された途端、会場内は爆笑の渦に包まれたという逸話が残るほどである[6][10]。
Whittington 1975 では、オパビニアはどの動物門にも属さず、節足動物と環形動物の体節制をもつ原始的な系統群の種類ではないかと解釈された[6](節足動物と環形動物は当時ではまだ一般に近縁扱いされ、お互いに別系統だと判明したのは21世紀以降である。節足動物#他の動物門との関係性を参照[44])。その難解さはスティーヴン・ジェイ・グールド(Stephen Jay Gould)著の『ワンダフルライフ』(1989)で更に強調され、同じくカンブリア紀のアノマロカリスなどと共に、現生の動物門に分類不可能な「プロブレマティカ」(未詳化石)で、バージェス動物群の「奇妙奇天烈動物」(Weird Wonders)の代表例として取り上げられた[12]。
基盤的な節足動物(1980年代後期~21世紀)
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| 系統解析に支持されるオパビニアの系統的位置[5] †:絶滅群、青枠:基盤的な節足動物、*:葉足動物 |








しかし前述の見解に対して専門家からの反発が強く、アノマロカリスと共に、オパビニアはそこまで言うほど難解ではなく、節足動物に含める可能性が多くの文献記載に示された[7][34][8][3][21]。特に1990年代以降では、新たな発見と系統分類学(特に分岐分類学)の発展により、これらの古生物の類縁関係についての考察が飛躍的に進んでいた[10]。
Bergström 1986 の再記載をはじめとして、オパビニアとアノマロカリスの共通点(特化した前部付属肢・眼柄に突出した複眼・胴部の鰭と鰓・尾扇など)が注目される同時に、今まで思った以上に節足動物的であることも徐々に判明した[7][25]。オパビニアの吻は一見して異様だが、先端の"ハサミ"は(Whittington 1975 [6]のような縦開きではなく)横開きで左右対称の構造体であり、明らかに融合した1対の付属肢である[7][25]。このような付属肢の融合に無理はなく、むしろ節足動物で一般に見られる特化様式である[7]。鰓の部分もアノマロカリスと同様に一連の葉状の附属体であると判明し[7]、これは節足動物の外葉によく似ている[8][13][9][29]。更に、両者の特化した消化腺[31]と眼柄に突出した複眼・オパビニアの後ろ向きの口・アノマロカリスの関節肢化した前部付属肢も節足動物的な特徴である[7][25][2]。一方で、オパビニアとアノマロカリスの放射状の口器と柔軟な胴部は(付属肢由来の口器と硬質の外骨格を胴部にもつ)一般的な節足動物とは明らかに異なるため、これらの古生物は、少なくとも現生の節足動物の系統群に属さない「独特な節足動物」であることも同時に示された[7][25][3]。
1990年代では、ケリグマケラとパンブデルリオンが見つかり、この2つの古生物はオパビニアとアノマロカリスに似た特徴(前部付属肢・鰭・鰓・消化腺)だけでなく、葉足動物に似た性質(柔軟な表皮・環形の筋・葉足・放射状の口器)も出揃った中間型生物(ミッシングリンク)であり、前述の節足動物らしからぬ性質は、汎節足動物や脱皮動物の起源まで遡れる、葉足動物との共通祖先から受け継いた祖先形質であることを示していた[34][8][28][21]。こうしてオパビニアとこれらの古生物は、胴部の外骨格を進化する以前の、節足動物の絶滅した初期系統(ステムグループ)に属する「基盤的な節足動物」である説と、節足動物は(有爪動物・緩歩動物と同様に)葉足動物から進化した動物群である説が台頭し始めた[34][8][28][21]。


アノマロカリスは、後に多くの近縁(ペイトイア、フルディア、アンプレクトベルアなど)をもつことが分かり、ラディオドンタ類(Radiodonta、旧称アノマロカリス類)の一員としてまとめられるようになった[3]。オパビニアはラディオドンタ類ではないが、オパビニア類(オパビニア科、後述参照)の一員としてラディオドンタ類と共に、基盤的な節足動物の中でケリグマケラとパンブデルリオンより派生的で、残りの「真正」の節足動物(真節足動物)に最も近縁の群と考えられるようになった[8][21]。この系統関係は、21世紀以降も多くの系統解析[32][45][46][47][48][49][50][51][52][30][53][54][55][56][22][57][58][59][60][61][62][63][64][65][66][67][68][69][23][70][71][72][5][24]と新たな発見[13][9](例えば中間型生物キリンシア[23])に広く支持され続けており、オパビニアとこれらの古生物(恐蟹類[3][20])は、節足動物の起源に重要な情報を与えたものとして一般に知られるようになった[9][10][2][14][15][16][17]。
近縁

オパビニアはオパビニア科(Opabiniidae、オパビニア類 opabiniid とも)の模式属(タイプ属)である[4]。オパビニア科は、1本の吻に癒合した前部付属肢・柔軟だが明確した境目をもつ胴節、少なくとも鰭の一部を覆い被さる鰓・複数対の尾鰭・1対の短い棘状の尾毛を兼ね備えることを特徴とし、同じ基盤的な節足動物とされるラディオドンタ類、パンブデルリオンとケリグマケラから区別される[5]。2022年現在、確実にオパビニア科に含める属はオパビニアとユタウロラ[73](Utaurora)のみである[5]。ユタウロラは胴節の背面まで覆い被さる鰓と7対以上の尾鰭によりオパビニア(鰓は胴節を覆わず、尾鰭3対)から区別される[5]。
→詳細は「オパビニア科」を参照
Collins 1996 をはじめとして、オパビニア科はラディオドンタ類などと共に 恐蟹類(dinocaridids, Dinocaridida)という、基盤的な節足動物の分類群(綱)に含まれるようになった[3][51][26][5]。その中でオパビニアは、ケリグマケラやパンブデルリオンと共に便宜的に「gilled lobopodians」("鰓のある葉足動物")と総称されることもある[8][2]。しかし、オパビニアに関しては葉足の有無が議論的で(後述参照)、ケリグマケラとパンブデルリオンほど顕著な葉足動物的性質もないため、葉足動物扱いされない場合がほとんどである[13][74]。
→詳細は「葉足動物 § 広義の_"葉足動物"」を参照
下位分類
1970年代[43][6]以降、オパビニア(オパビニア属 Opabinia)の構成種として認められるのは、2022年現在では未だに模式種(タイプ種)であるオパビニア・レガリス(Opabinia regalis)のみである[5]。原記載(Walcott 1912)では模式種の他に Opabinia? media も記載された[4]が、これは Hutchinson 1930 に O. regalis の幼生と考えされ、すなわち O. regalis のジュニアシノニム(無効異名)とされた[42]。Simonetta 1970 と Whittington 1975 では、O.? media の基準となった化石標本は独立種として判断しにくいほど産状が悪いと示され、O.? media は無効名とされた[43][6]。シベリアで見つかり、Miroshnikov & Krawzov 1960 に本属の種 Opabinia norilica として復元された悪い産状の化石標本もあった[19]が、これは別の節足動物の見間違いだった可能性が高く、Whittington 1975 に本属の種として認められなかった[6]。
鰭・鰓の構造と脚の有無

A: Whittington 1975 [6]
B: Bergström 1986 [7]
C: Budd 1996 [8]
D: Zhang & Briggs 2007 [13]
E: Budd & Daley 2011 [9]

オパビニアの様々な特徴の中でも、鰭と鰓の構造は更新頻度が最も高い部分であり、再記載(Whittington 1975[6]、Bergström 1986[7]、Budd 1996[8]、Zhang & Briggs 2007[13]、Budd & Daley 2011[9])ごとに解釈が変わる[10]。また、化石標本の各胴節と鰭の間には、やや長い三角型の痕跡が1対ずつ並んでおり、この部分の正体に関しては、主に「消化管の枝」と「柔らかい脚(葉足)」という2つの仮説が繰り広げられてきた[10]。なお、これらの部分の解釈の違いは、オパビニアの上述の分類学的位置にさほどの影響を与えていない[10][22](Whittington 1975 以外ではいずれも結論的にオパビニアの節足動物的性質を支持し[7][8][13][9]、脚の有無は系統解析の結果を影響しなかった[22])。
Whittington 1975 では、第1胴節に鰓はなく、鰓は鰭から完全に独立した出っ張りの筋とされた[6]。三角型の痕跡は内部構造だと思われ、消化管の枝、もしくは鰓に繋がる循環系の一部だと解釈された[6]。
Bergström 1986 では、鰓は独立した出っ張りの筋ではなく、多数の細長い葉状の附属体でできている、鰭に沿って連結する櫛状の構造体と判明した[7]。しかし、鰭が背板の出っ張り(pleurae)と解釈され、鰓はその腹面に連結すると考えられた[7]。

Budd 1996 では、鰓は鰭の背面に連結すると判明した同時に、鰓の連結部は鰭の前縁直後にあり、第1胴節は Whittington 1975 のように鰓はないとされた[8]。三角型の痕跡は内臓としてオパビニアの胴部に納めるには長すぎであると指摘され、側面に保存された化石標本ではその付け根も(側面にある)鰭の連結部からかけ離れてた腹面にあり、鰭から独立したことが示唆される[8]。これは葉足の体腔由来の痕跡とされ、すなわち各胴節の腹側には1対の短い葉足があり、加えて各葉足の先端に爪があることも推測された[8]。

Zhang & Briggs 2007 では、鰭の後縁は大きな切り込みで、そこに鰓が占めていると解釈された[13]。三角形の痕跡の元素組成が元素マッピング(elemental mapping)で分析され、消化管と同じ成分をもつことが判明した[13]。これを基に、Budd 1996 に提唱された葉足と爪の存在が否定され、この部分は(付け根が両腹面にある)鰭まで差し込んだ消化管の枝だと解釈された[13]。

既存と新たな化石標本に基づいた Budd & Daley 2011 の再検証では、葉足の存在を断言しないものの、その可能性を支持し、Budd 1996 の解釈をほぼ認めつつ、Zhang & Briggs 2007 の多くの判断を否定する(爪の存在は否定的であることのみ認められる)幾つかの証拠が追記された[9]。良好に保存されたオパビニアの鰭から完全の後縁が見られ、Zhang & Briggs 2007 に提唱される切り込みは存在しないことが証明された[9]。鰭の鰓との連結部は Budd 1996 の解釈より少し後ろに位置し、第1胴節の鰭にも鰓があることも判明した[9]。消化管と三角形の痕跡にある同様の元素組成は、吻など他の部位にも見つかり、消化管に限られるものではない[9]。消化管の枝に該当する部位は、実はその痕跡とは全く別の部位である、数対の丸い消化腺であることも明らかにされた[9]。葉足動物であるアイシェアイアの化石標本に見られる葉足の体腔痕跡とオパビニアの三角形の痕跡の類似性も示され、更に一部のオパビニアの化石標本からは、僅かであるものの、葉足の外皮組織に似た、環形の筋に分かれる構造も発見された[9]。
脚注
関連項目
外部リンク
Wikiwand - on
Seamless Wikipedia browsing. On steroids.