Top Qs
Timeline
Obrolan
Perspektif
Arkea
domain dari mikroorganisme prokariotik Dari Wikipedia, ensiklopedia bebas

Remove ads
Arkea atau Archaea adalah domain mikroorganisme bersel satu. Arkea tergolong prokariota, kelompok mikrob yang tidak memiliki inti sel dan organel yang dibatasi membran. Awalnya, arkea diklasifikasikan sebagai bakteri dan diberi nama archaebacteria (di kerajaan Archaebacteria), tetapi klasifikasi ini tak lagi digunakan.

Sel arkea memiliki sifat unik yang memisahkan arkea dari dua domain kehidupan yang lain, yaitu Bacteria dan Eukaryota. Arkea dibagi lagi menjadi beberapa filum. Klasifikasinya sulit dilakukan karena mayoritas arkea belum diisolasi di laboratorium dan hanya dideteksi urutan gennya dari sampel yang diambil dari lingkungan.
Arkea dan bakteri umumnya memiliki ukuran dan bentuk yang serupa, meskipun beberapa arkea memiliki bentuk yang sangat aneh, misalnya sel-sel datar dan berbentuk persegi seperti Haloquadratum walsbyi. Meskipun terdapat kesamaan morfologi antara arkea dengan bakteri, arkea memiliki gen-gen dan beberapa jalur metabolik yang lebih serupa dengan eukariota, terutama enzim yang terlibat dalam transkripsi dan translasi. Salah satu aspek biokimia arkea yang unik adalah ketergantungan arkea pada lipid eter di membran sel mereka. Arkea menggunakan lebih banyak sumber energi dibandingkan eukariota, yang berkisar dari senyawa organik seperti gula, amonia, ion logam, atau bahkan gas hidrogen. Arkea yang toleran terhadap garam (Haloarchaea) menggunakan sinar matahari sebagai sumber energi, sementara spesies arkea lain memfiksasi karbon; namun, tidak seperti tumbuhan dan sianobakteri, tidak ada arkea yang melakukan kedua cara tersebut. Arkea bereproduksi secara aseksual dengan pembelahan biner, fragmentasi, atau tunas. Tidak seperti bakteri dan eukariota, tidak ada spesies arkea yang diketahui membentuk endospora. Arkea pertama yang diamati merupakan ekstremofil yang hidup di lingkungan ekstrem, seperti mata air panas dan danau garam tanpa organisme lain. Meningkatnya kemampuan alat deteksi molekuler memungkinkan penemuan arkea di hampir semua habitat, termasuk tanah, lautan, dan rawa. Jumlah mereka sangat banyak di lautan, dan arkea dalam bentuk plankton mungkin merupakan salah satu kelompok organisme yang paling melimpah di planet ini.
Arkea berperan besar dalam kehidupan di Bumi. Mereka merupakan bagian dari mikrobiota bagi semua organisme. Sebagai mikrobiota manusia, arkea berperan penting di usus besar, rongga mulut, dan kulit. Keragaman morfologis, metabolik, dan geografis memungkinkan mereka memiliki peran ekologis yang luas, misalnya pada fiksasi karbon dalam siklus karbon, peran dalam siklus nitrogen, perputaran senyawa organik, dan memelihara komunitas simbiosis dan sintrofi mikrob. Belum ditemukan contoh yang jelas untuk arkea yang bersifat patogenik atau parasitik. Sebaliknya, arkea sering kali bersifat mutualis atau komensal, misalnya metanogen yang hidup di saluran pencernaan manusia dan ruminansia, tempat mereka hidup dalam jumlah besar dan membantu pencernaan. Metanogen digunakan dalam produksi biogas dan pengolahan limbah. Sementara itu, enzim dari arkea ekstremofil yang dapat bertahan pada suhu tinggi dan pelarut organik dimanfaatkan dalam bioteknologi.
Remove ads
Klasifikasi
Ringkasan
Perspektif
Konsep awal
.jpg)
Pada hampir sepanjang abad ke-20, prokariota dianggap sebagai satu kelompok organisme dan diklasifikasikan berdasarkan sifat biokimia, morfologi, dan metabolisme mereka. Para ahli mikrobiologi mencoba mengklasifikasikan mikroorganisme berdasarkan struktur dinding sel, bentuk, dan zat-zat yang mereka konsumsi.[6] Pada tahun 1965, Linus Pauling dan Emile Zuckerkandl[7] mengusulkan untuk menggunakan urutan gen dalam berbagai prokariota untuk mengetahui hubungan mereka satu sama lain. Pendekatan ini, yang dikenal sebagai filogenetik, adalah metode utama yang digunakan saat ini.[8]
Arkea (saat itu hanya metanogen yang diketahui) pertama kali diklasifikasikan sebagai kelompok yang terpisah dari bakteri pada pohon filogenetik berdasarkan urutan gen RNA ribosomal (rRNA) mereka oleh Carl Woese dan George E. Fox pada tahun 1977.[9] Awalnya, kedua kelompok tersebut masing-masing diberi nama Archaebacteria dan Eubacteria, dan diperlakukan sebagai kerajaan atau subkerajaan, yang oleh Woese dan Fox disebut Urkingdoms. Namun, klasifikasi ini tak lagi digunakan.[10] Woese dan Fox berpendapat bahwa Archaebacteria secara fundamental berbeda dari kehidupan yang lain: (1) ketiadaan peptidoglikan pada dinding sel mereka; (2) adanya dua koenzim yang tidak biasa; dan (3) hasil pengurutan gen RNA ribosomal 16S. Untuk menekankan perbedaan-perbedaan ini, Woese, Otto Kandler, dan Mark Wheelis kemudian mengusulkan sistem pengelompokan organisme yang baru dengan tiga domain terpisah: Eukarya, Bacteria, dan Archaea,[1] yang sekarang dikenal sebagai "Revolusi Woesian".[11]
Kata arkea berasal dari bahasa Yunani Kuno ἀρχαῖα, yang berarti "hal-hal kuno",[12] karena wakil pertama dari domain Archaea adalah metanogen dan diasumsikan bahwa metabolisme mereka mencerminkan suasana primitif Bumi dan mereka merupakan organisme kuno. Untuk waktu yang lama, arkea terlihat sebagai ekstremofil yang hanya ada di habitat ekstrem seperti mata air panas dan danau garam. Namun, seiring bertambahnya habitat baru yang dipelajari, semakin banyak arkea yang ditemukan. Mikrob-mikrob halofilik ekstrem[13] dan hipertermofilik[14] juga termasuk dalam Archaea. Pada akhir abad ke-20, arkea juga telah diidentifikasi di lingkungan nonekstrem. Saat ini, mereka dikenal sebagai kelompok organisme yang besar dan beragam yang tersebar luas di alam.[15] Arkea mendapatkan apresiasi baru mengenai peranan dan keberadaan mereka sejak reaksi berantai polimerase (PCR) digunakan untuk mendeteksi prokariota dari sampel lingkungan (seperti air atau tanah) dengan melipatgandakan gen ribosom mereka. Hal ini memungkinkan deteksi dan identifikasi organisme yang belum dibudidayakan di laboratorium.[16][17]
Klasifikasi

Klasifikasi arkea, dan prokariota pada umumnya, adalah bidang yang bergerak cepat dan diperdebatkan. Sistem klasifikasi saat ini bertujuan untuk mengatur arkea ke dalam kelompok organisme yang memiliki karakteristik struktural dan nenek moyang yang sama.[18] Klasifikasi ini sangat bergantung pada pengurutan gen RNA ribosomal untuk mengungkapkan hubungan di antara berbagai organisme, yang dikenal sebagai filogenetik molekuler.[19] Sebagian besar spesies arkea yang dapat dikultur dan diteliti dengan baik adalah anggota dari dua filum utama, Euryarchaeota dan Crenarchaeota. Kelompok-kelompok lain dibuat secara tentatif. Misalnya, spesies aneh Nanoarchaeum equitans, yang ditemukan pada tahun 2003 dan telah diberikan filum sendiri, Nanoarchaeota.[20] Korarchaeota juga telah diusulkan sebagai filum baru yang berisi sekelompok kecil spesies termofilik yang berbagi karakteristik dengan kedua filum utama, tetapi lebih berkerabat dekat dengan Crenarchaeota.[21][22] Spesies arkea lain ternyata berkerabat jauh dengan salah satu dari kelompok ini, seperti Archaeal Richmond Mine acidophilic nanoorganisms (ARMAN) yang ditemukan pada tahun 2006[23] dan merupakan salah satu organisme terkecil yang diketahui.[24]
Sebuah superfilum, yang disebut TACK, yang mencakup Thaumarchaeota, Aigarchaeota, Crenarchaeota, dan Korarchaeota, diusulkan pada 2011 dan mungkin terkait dengan organisme yang menjadi moyang eukariota.[25] Pada tahun 2017, superfilum Asgard telah diajukan sebagai kelompok yang lebih dekat dengan eukariota dan merupakan kelompok saudari dari TACK.[26]
Kladogram
Berikut ini adalah kladogram arkea berdasarkan Tom A. Williams dkk. (2017)[27] serta Castelle dan Banfield (2018)[28] (DPANN):
Konsep spesies
Klasifikasi spesies-spesies arkea juga kontroversial. Biologi mendefinisikan spesies sebagai sekelompok organisme yang saling terkait. Kriteria perkembangbiakan eksklusif yang diketahui secara umum (organisme dalam satu spesies dapat berkembang biak dengan satu sama lain tetapi tidak dengan yang lain) tidak membantu karena arkea bereproduksi secara aseksual.[32]
Arkea menunjukkan tingkat transfer gen horizontal yang tinggi di antara garis keturunan mereka. Beberapa peneliti menyarankan agar sejumlah individu dikelompokkan ke dalam populasi-populasi yang menyerupai spesies, yang genom-genomnya sangat mirip dan jarang mentransfer gen atau menerima transfer gen dari sel lain yang genomnya kurang terkait, seperti dalam genus Ferroplasma.[33] Di sisi lain, studi terhadap Halorubrum menemukan adanya transfer genetik yang signifikan ke/dari populasi yang kurang terkait, sehingga penerapan kriteria ini terbatas.[34] Kekhawatiran kedua adalah sejauh mana sebutan spesies memiliki arti praktis.[35]
Pengetahuan terkini tentang keanekaragaman genetik masih tidak lengkap dan jumlah spesies arkea tidak dapat diperkirakan dengan akurat.[19] Perkiraan jumlah filum berkisar antara 18–23, tetapi hanya delapan filum yang memiliki perwakilan yang telah dikultur di laboratorium dan dipelajari secara langsung. Banyak kelompok-kelompok arkea dihipotesiskan hanya dari urutan rRNA tunggal, yang menunjukkan bahwa perbedaan di antara organisme ini tetap tidak jelas.[36] Bakteri juga mengandung banyak mikrob yang belum dikultur sehingga karakterisasi mereka juga serupa dengan arkea.[37]
Remove ads
Asal-usul dan evolusi
Ringkasan
Perspektif
Umur Bumi diperkirakan sekitar 4,54 miliar tahun.[38][39][40] Bukti ilmiah menunjukkan bahwa kehidupan di Bumi dimulai setidaknya 3,5 miliar tahun yang lalu.[41][42] Bukti-bukti awal kehidupan di Bumi adalah grafit yang bersifat biogenik pada batuan metasedimentari berumur 3,7 miliar tahun yang ditemukan di Greenland Barat[43] dan fosil tikar mikrob yang ditemukan di batu pasir setua 3,48 miliar tahun di Australia Barat.[44][45] Pada tahun 2015, kemungkinan "sisa-sisa kehidupan biotik" ditemukan di batuan berumur 4,1 miliar tahun di Australia Barat.[46][47]
Meskipun fosil sel prokariota kemungkinan berasal dari hampir 3,5 miliar tahun yang lalu, sebagian besar prokariota tidak memiliki morfologi yang dapat dibedakan, dan bentuk fosil tidak dapat digunakan untuk mengidentifikasi mereka sebagai arkea.[48] Sebaliknya, fosil kimia lipid yang unik menjadi lebih informatif karena senyawa tersebut tidak ada pada organisme lain.[49] Beberapa publikasi menyatakan bahwa sisa-sisa lipid arkea atau eukariota yang hadir dalam batu serpih berasal dari 2,7 miliar tahun yang lalu;[50] meskipun data tersebut dipertanyakan.[51] Lipid tersebut juga terdeteksi di batuan yang bahkan lebih tua dari batuan di Greenland Barat. Jejak tertua berasal dari distrik Isua, Greenland, yang termasuk sedimen tertua yang dikenal di Bumi yang terbentuk 3,8 miliar tahun yang lalu.[52] Silsilah arkea mungkin menjadi garis keturunan yang paling kuno yang ada di Bumi.[53]
Woese berpendapat bahwa bakteri, arkea, dan eukariota mewakili garis keturunan yang terpisah yang bercabang sejak awal dari koloni leluhur semua organisme.[54][55] Salah satu kemungkinannya[55][56] adalah bahwa percabangan ini terjadi sebelum evolusi sel, saat tidak adanya membran sel memungkinkan transfer gen lateral yang tak terbatas, dan bahwa nenek moyang dari tiga domain tersebut muncul dari fiksasi subset-subset gen yang spesifik.[55][56] Ada kemungkinan bahwa nenek moyang terakhir dari bakteri dan arkea adalah termofil, sehingga suhu yang lebih rendah mungkin saja merupakan "lingkungan yang ekstrem" bagi arkea, dan organisme-organisme yang hidup di lingkungan dingin baru muncul belakangan.[57] Karena kekerabatan Archaea dan Bacteria lebih berjauhan satu sama lain dibandingkan antara mereka dengan eukariota, istilah prokariota mungkin menunjukkan kesamaan yang salah antara arkea dan bakteri.[58]
Perbandingan dengan domain lain
Tabel berikut membandingkan beberapa karakteristik utama dari tiga domain, untuk menggambarkan persamaan dan perbedaan mereka.[59]
Archaea dipisahkan sebagai domain ketiga karena memiliki perbedaan besar dalam struktur RNA ribosomal mereka. Molekul RNA tertentu yang dikenal sebagai RNA ribosomal 16S (biasanya disingkat menjadi 16s rRNA) adalah kunci penting dalam produksi protein di semua organisme. Karena fungsi ini sangat penting dalam kehidupan, organisme yang mengalami mutasi 16s rRNA tidak mungkin bertahan hidup sehingga struktur nukleotida ini menjadi sangat stabil (tetapi tidak absolut) selama beberapa generasi. Ukuran 16s rRNA juga cukup besar untuk menyimpan informasi spesifik pada organisme tertentu, tetapi cukup kecil untuk diurutkan dan dibandingkan dalam waktu singkat. Pada tahun 1977, Carl Woese, seorang ahli mikrobiologi yang mempelajari urutan genetik organisme, mengembangkan metode perbandingan baru yang melibatkan pemecahan RNA menjadi fragmen-fragmen yang bisa diurutkan dan dibandingkan dengan fragmen lain dari organisme lain.[9] Semakin mirip pola fragmen tersebut di antara spesies-spesies, hubungan kekerabatannya menjadi lebih dekat.[62]
Woese menggunakan metode perbandingan rRNA barunya untuk mengelompokkan dan memisahkan organisme yang berbeda. Ia mengurutkan berbagai spesies dan menemukan bahwa sekelompok metanogen dengan rRNA ternyata sangat berbeda dibandingkan prokariota atau eukariota yang telah dikenal.[9] Metanogen ini jauh lebih mirip satu sama lain dibandingkan dengan organisme lain yang pernah diurutkan, sehingga Woese mengusulkan domain baru, yaitu Archaea.[9] Eksperimen Woese menunjukkan bahwa arkea lebih mirip dengan eukariota alih-alih prokariota, meskipun struktur mereka lebih mirip dengan prokariota.[63] Hal ini menyimpulkan bahwa Archaea dan Eukarya memiliki nenek moyang bersama yang lebih baru dibandingkan dengan Eukarya dan Bacteria.[63] Perkembangan inti sel terjadi setelah percabangan antara bakteri dan nenek moyang bersama ini.[63]
Salah satu sifat arkea yang unik adalah penggunaan lipid dengan ikatan eter secara berlimpah di membran sel mereka. Secara kimiawi, ikatan eter lebih stabil dari ikatan ester yang ditemukan dalam Bacteria dan Eukarya, yang mungkin menjadi faktor yang berkontribusi terhadap kemampuan banyak arkea untuk bertahan di lingkungan ekstrem, yang memberikan stres berat pada membran sel, seperti panas ekstrem dan salinitas. Analisis komparatif genom arkea juga telah mengidentifikasi beberapa indel khusus yang terkonservasi secara molekuler dan protein tertentu yang secara unik ditemukan baik di semua arkea maupun ditemukan di beberapa kelompok-kelompok utama dalam arkea .[64][65][66] Sifat lain yang unik dari arkea adalah bahwa tidak ada organisme lain yang dikenal mampu melakukan metanogenesis (produksi metana melalui metabolisme). Arkea metanogen memainkan peran penting dalam ekosistem bersama dengan organisme yang memperoleh energi dari oksidasi metana (kebanyakan dari mereka adalah bakteri) karena mereka sering menjadi sumber utama metana di lingkungan tersebut dan dapat berperan sebagai produsen primer. Metanogen juga memainkan peran penting dalam siklus karbon, memecah karbon organik menjadi metana, yang juga merupakan gas rumah kaca utama.[67]
Hubungan dengan bakteri
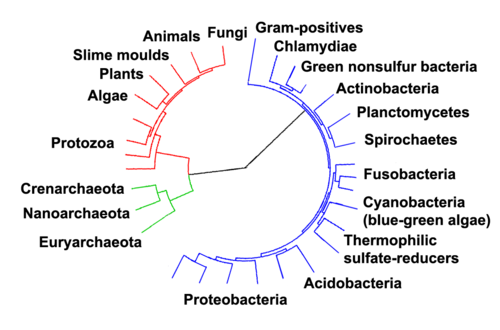
Hubungan di antara tiga domain sangat penting untuk memahami asal-usul kehidupan. Sebagian besar lintasan metabolisme (yang merupakan objek dari sebagian besar gen suatu organisme) sama-sama dimiliki Archaea dan Bacteria, sementara sebagian besar gen yang terlibat dalam ekspresi genom sama-sama dimiliki Archaea dan Eukarya.[69] Di antara prokariota, struktur sel arkea paling mirip dengan bakteri Gram-positif, terutama karena keduanya memiliki lipida dwilapis tunggal[70] dan biasanya mengandung sakulus tebal (eksoskeleton) dari berbagai komposisi kimia.[71] Pada beberapa pohon filogenetik yang didasarkan pada urutan gen-gen (protein-protein) dari homolog prokariota, homolog arkea lebih berhubungan dekat dengan homolog bakteri Gram-positif.[70] Arkea dan bakteri Gram-positif juga berbagi indel yang dilestarikan dalam sejumlah protein penting, seperti Hsp70 dan glutamin sintetase I;[70][72] namun, ilmuwan menafsirkan filogeni dari gen-gen ini sebagai sarana untuk mentransfer gen interdomain,[73][74] dan mungkin tidak mencerminkan hubungan kekerabatan organisme.[75]
Ada pendapat bahwa arkea berevolusi dari bakteri Gram-positif sebagai respons terhadap tekanan seleksi antibiotik.[70][72][76] Pandangan ini diajukan berdasarkan pengamatan bahwa arkea tahan terhadap berbagai antibiotik yang terutama dibuat untuk bakteri Gram-positif,[70][72] dan bahwa antibiotik-antibiotik ini terutama bekerja pada gen yang membedakan arkea dari bakteri. Pendapat ini menyatakan bahwa tekanan selektif terhadap resistansi yang dihasilkan oleh antibiotik Gram-positif akhirnya cukup untuk menyebabkan perubahan yang luas pada banyak gen target antibiotik, dan bahwa galur ini mewakili nenek moyang arkea masa kini.[76] Evolusi arkea dalam menanggapi pemilihan antibiotik, atau tekanan selektif kompetitif lain, juga bisa menjelaskan adaptasi mereka terhadap lingkungan yang ekstrem (seperti suhu tinggi atau keasaman) sebagai hasil dari pencarian relung kosong untuk melarikan diri dari organisme yang memproduksi antibiotik;[76][77] Cavalier-Smith juga membuat usulan yang sama.[78] Pendapat ini juga didukung oleh studi lainnya yang menyelidiki hubungan protein struktural[79] dan studi yang menunjukkan bahwa bakteri Gram-positif mungkin merupakan awal garis keturunan yang bercabang paling awal dalam prokariota.[80]
Hubungan dengan eukariota
Hubungan evolusi antara arkea dan eukariota masih belum jelas. Selain dari kesamaan dalam struktur dan fungsi sel yang dibahas di bawah, banyak pohon genetik menyatukan keduanya dalam satu kelompok.[81]
Faktor-faktor yang memperrumit hubungan mereka termasuk klaim bahwa hubungan antara eukariota dan filum arkea Crenarchaeota lebih dekat daripada hubungan antara filum Crenarchaeota dan Euryarchaeota,[82] dan adanya gen-seperti-arkea pada bakteri tertentu, seperti pada Thermotoga maritima, dari transfer gen horizontal.[83] Hipotesis standar menyatakan bahwa nenek moyang eukariota berpisah dari Archaea sejak awal,[84][85] dan bahwa eukariota muncul melalui fusi antara arkea dan bakteri, yang menjadi inti sel dan sitoplasma; hal ini menjelaskan berbagai kesamaan genetik tetapi kesulitan dalam menjelaskan struktur sel.[82] Sebuah hipotesis alternatif, hipotesis eosit, berpendapat bahwa Eukaryota muncul relatif terlambat dari Archaea.[86]
Sebuah garis keturunan arkea yang ditemukan pada 2015, Lokiarchaeum (dari filum baru Lokiarchaeota), yang dinamai berdasarkan sebuah ventilasi hidrotermal yang disebut Kastel Loki di Samudra Arktik, ternyata merupakan kelompok yang paling berkerabat dekat dengan eukariota. Organisme ini disebut sebagai organisme peralihan antara prokariota dan eukariota.[87][88]
Beberapa filum saudara dari "Lokiarchaeota" telah ditemukan ("Thorarchaeota", "Odinarchaeota", "Heimdallarchaeota"), yang semuanya membentuk sebuah supergrup Asgard yang baru diajukan, yang mungkin merupakan takson saudara dari Proteoarchaeota.[3][26][89] Rincian dari hubungan anggota Asgard dan eukariota masih dalam pertimbangan,[90] meskipun, pada Januari 2020, para ilmuwan melaporkan bahwa Candidatus Prometheoarchaeum syntrophicum, sejenis arkea Asgard, mungkin merupakan penghubung antara mikroorganisme prokariota yang sederhana dan mikroorganisme eukariota yang kompleks sekitar dua miliar tahun yang lalu.[91][92]
Remove ads
Morfologi
Ringkasan
Perspektif
Satu individu arkea memiliki diameter yang berkisar dari 0,1 mikrometer (μm) sampai lebih dari 15 μm, yang hadir dalam berbagai bentuk, umumnya sebagai bola, batang, spiral, atau piring.[93] Morfologi lain ditemukan pada filum Crenarchaeota, termasuk sel berlobus yang bentuknya tidak teratur pada genus Sulfolobus, filamen seperti-jarum yang diameternya kurang dari setengah mikrometer pada genus Thermofilum, serta batang yang bentuknya hampir persegi panjang sempurna pada Thermoproteus dan Pyrobaculum.[94] Haloquadratum walsbyi adalah arkea dengan sel berbentuk persegi yang datar, yang hidup di kolam hipersalin.[95][96] Bentuk yang tidak biasa ini mungkin dipertahankan dengan baik oleh dinding sel dan sitoskeleton prokariotik mereka. Protein yang terkait dengan komponen sitoskeleton organisme lain ditemukan di arkea,[97] dan filamen-filamen terbentuk dalam sel mereka,[98] tetapi berbeda dengan organisme lain, struktur seluler ini kurang dipahami.[99] Dalam Thermoplasma dan Ferroplasma, ketiadaan dinding sel berarti bahwa sel memiliki bentuk yang tidak teratur, dan dapat menyerupai ameba.[100]
Beberapa spesies arkea membentuk agregat atau filamen sel yang panjangnya hingga 200 μm.[93] Organisme-organisme ini bisa membentuk tonjolan dalam biofilm.[101] Khususnya, agregat sel-sel Thermococcus coalescens menyatu bersama dalam kultur sehingga membentuk sel-sel raksasa tunggal.[102] Genus Pyrodictium menghasilkan koloni multisel rumit yang melibatkan tabung berongga panjang dan tipis yang disebut kanula yang menonjol dari permukaan sel dan menghubungkan mereka ke dalam aglomerasi padat seperti-semak.[103] Fungsi dari kanula ini tidak disepakati, tetapi mereka dapat memungkinkan komunikasi atau pertukaran nutrisi dengan tetangganya.[104] Koloni multispesies juga ditemukan, seperti komunitas "untaian mutiara" yang ditemukan pada tahun 2001 di sebuah rawa di Jerman. Koloni bulat keputihan dari spesies baru Euryarchaeota terletak di sepanjang filamen tipis yang panjangnya dapat mencapai 15 sentimeter (5,9 inci); filamen ini terbuat dari spesies bakteri tertentu.[105]
Remove ads
Struktur, perkembangan komposisi, dan operasi
Ringkasan
Perspektif
Arkea dan bakteri memiliki struktur sel yang umumnya sama, tetapi komposisi dan organisasi sel membedakan arkea dengan bakteri. Seperti bakteri, arkea tidak memiliki membran interior dan organel.[58] Membran sel arkea, juga seperti bakteri, biasanya dibatasi oleh dinding sel dan mereka "berenang" menggunakan satu flagela atau lebih.[106] Secara struktural, arkea paling mirip dengan bakteri Gram-positif. Kebanyakan arkea memiliki membran plasma dan dinding sel tunggal, tanpa ruang periplasmik; pengecualian untuk aturan umum ini adalah Ignicoccus, yang memiliki sebuah periplasma sangat besar yang berisi vesikel bermembran dan tertutup oleh membran luar.[107]
Dinding sel dan flagela
Kebanyakan arkea (kecuali Thermoplasma dan Ferroplasma) memiliki dinding sel.[100] Mayoritas dinding sel arkea dirakit dari protein lapisan permukaan, yang membentuk lapisan-S.[108] Lapisan-S adalah susunan molekul protein yang kaku yang menutupi bagian luar sel (seperti zirah rantai).[109] Lapisan ini memberi perlindungan kimia dan fisik, serta dapat mencegah makromolekul mengalami kontak dengan membran sel.[110] Tidak seperti bakteri, arkea tidak memiliki peptidoglikan di dinding sel mereka.[111] Methanobacteriales memiliki dinding sel yang mengandung pseudopeptidoglikan, yang menyerupai peptidoglikan eubacteria dalam morfologi, fungsi, dan struktur fisik, tetapi pseudopeptidoglikan berbeda dalam struktur kimia; tidak memiliki asam amino-D dan asam N-asetilmuramat.[110]
Flagela arkea beroperasi seperti flagela bakteri—tangkai panjangnya digerakkan oleh motor yang berputar di pangkalnya. Motor ini didukung oleh gradien proton yang melintasi membran. Namun, flagela arkea berbeda, terutama dalam komposisi dan pengembangan.[106] Kedua jenis flagela (milik bakteri dan arkea) berevolusi dari nenek moyang yang berbeda. Flagela bakteri memiliki satu nenek moyang dengan sistem sekresi tipe III,[112][113] sementara flagela arkea tampaknya berevolusi dari pili bakteri tipe IV.[114] Berbeda dengan flagela bakteri yang berongga dan dirakit oleh subunit yang bergerak naik dari pori tengah ke ujung flagela, flagela arkea disintesis dengan menambahkan subunit di pangkalnya.[115]
Membran

Membran arkea terbuat dari molekul yang sangat berbeda dari molekul di organisme lain, yang menunjukkan bahwa arkea berkerabat jauh dari bakteri dan eukariota.[116] Pada semua organisme, membran sel terbuat dari molekul yang dikenal sebagai fosfolipid. Molekul-molekul ini memiliki bagian kutub yang larut dalam air ("kepala" fosfat) dan bagian nonpolar "berminyak" yang tidak larut dalam air (ekor lipid). Bagian yang berbeda ini dihubungkan oleh gugus gliserol. Dalam air, fosfolipid mengelompok, dengan kepala menghadap air dan ekor menjauhi air. Struktur utama dalam membran sel adalah lapisan ganda fosfolipid ini, yang disebut lipida dwilapis.[117]
Fosfolipid yang dimiliki arkea tidak biasa dalam empat hal:
- Bakteri dan eukariota memiliki membran yang terutama tersusun atas gliserol-lipid ester, sedangkan arkea memiliki membran yang tersusun atas gliserol-lipid eter.[118][119] Perbedaannya adalah jenis ikatan yang menggabungkan lipid ke bagian gliserol; dua jenis ikatan ditunjukkan dengan warna kuning pada gambar di sebelah kanan. Pada lipid ester, ikatan ini merupakan ikatan ester, sedangkan pada lipid eter ikatan ini merupakan ikatan eter. Ikatan eter secara kimiawi lebih tahan dari ikatan ester. Stabilitas ini mungkin membantu arkea bertahan pada suhu ekstrem dan lingkungan yang sangat asam atau basa.[120]
- Stereokimia dari bagian gliserol arkea berkebalikan dari yang ditemukan pada organisme lain. Bagian gliserol dapat terjadi dalam dua bentuk yang merupakan bayangan cermin satu sama lain, yang disebut bentuk tangan kanan dan tangan kiri; dalam kimia hal ini disebut enantiomer. Seperti tangan kanan yang tidak cocok dan sulit masuk ke dalam sarung tangan kiri, fosfolipid tangan kanan umumnya tidak dapat digunakan atau dibuat oleh enzim yang disesuaikan untuk bentuk kiri. Hal ini menunjukkan bahwa arkea menggunakan enzim yang sama sekali berbeda untuk sintesis fosfolipid dibandingkan bakteri dan eukariota. Enzim-enzim seperti ini dikembangkan sangat awal dalam sejarah kehidupan, yang menunjukkan percabangan awal dari dua domain lainnya.[116]
- Ekor lipid arkea secara kimiawi berbeda dari organisme lain. Lipid arkea didasarkan pada rantai isoprenoid panjang dengan banyak cabang samping, dan terkadang dengan cincin siklopropana atau sikloheksana.[121] Hal ini kontras dengan asam lemak yang ditemukan dalam membran organisme lain, yang memiliki rantai lurus tanpa cabang atau cincin. Meskipun isoprenoid memainkan peran penting dalam biokimia banyak organisme, hanya arkea yang menggunakannya untuk membuat fosfolipid. Rantai ini bercabang dapat membantu mencegah kebocoran membran arkea pada suhu tinggi.[122]
- Pada beberapa arkea, lapisan ganda lipid digantikan oleh sebuah lapisan tunggal. Akibatnya, arkea menyatukan dua ekor molekul fosfolipid independen menjadi molekul tunggal dengan dua kepala polar (bolaampifil); fusi ini dapat membuat membran mereka lebih kaku dan lebih mampu menahan lingkungan yang keras.[123] Sebagai contoh, lipid dalam Ferroplasma memiliki jenis lapisan ini, yang diduga untuk membantu kelangsungan hidup organisme ini di habitatnya yang sangat asam.[124]
Remove ads
Metabolisme
Ringkasan
Perspektif
Arkea menunjukkan berbagai macam reaksi kimia dalam metabolisme mereka dan menggunakan banyak sumber energi. Reaksi-reaksi ini diklasifikasikan ke dalam kelompok nutrisional, tergantung pada sumber-sumber energi dan karbon. Beberapa arkea memperoleh energi dari senyawa anorganik seperti belerang atau amoniak (mereka disebut litotrof). Arkea jenis ini termasuk penitrifkasi, metanogen, dan oksidator metana anaerob.[125] Dalam reaksi ini, satu senyawa memberikan elektron kepada senyawa yang lain (dalam reaksi redoks), melepaskan energi untuk bahan bakar aktivitas sel. Satu senyawa bertindak sebagai donor elektron dan satu sebagai akseptor elektron. Energi yang dilepaskan menghasilkan adenosina trifosfat (ATP) melalui kemiosmosis, dalam proses dasar yang sama yang terjadi di mitokondria sel eukariotik.[126]
Kelompok arkea lain menggunakan sinar matahari sebagai sumber energi (mereka disebut fototrof). Namun, fotosintesis yang menghasilkan oksigen tidak terjadi dalam organisme-organisme ini.[126] Banyak jalur metabolik dasar dimiliki bersama oleh semua bentuk kehidupan; misalnya, arkea menggunakan bentuk modifikasi dari glikolisis (jalur Entner–Doudoroff) serta salah satu dari siklus asam sitrat lengkap atau parsial.[127] Kesamaan dengan organisme lain ini mungkin mencerminkan bahwa jalur-jalur metabolik dasar tersebut memiliki asal-usul pada awal sejarah kehidupan dan tingkat efisiensi yang tinggi.[128]
Beberapa Euryarchaeota adalah metanogen yang hidup di lingkungan anaerobik, seperti rawa-rawa. Bentuk metabolisme ini berevolusi sejak awal, dan bahkan mungkin saja organisme yang pertama hidup bebas adalah metanogen.[129] Umumnya, reaksi melibatkan penggunaan karbon dioksida sebagai akseptor elektron untuk mengoksidasi hidrogen. Metanogenesis melibatkan berbagai koenzim yang unik untuk arkea ini, seperti koenzim M dan metanofuran.[130] Senyawa-senyawa organik lain seperti alkohol, asam asetat, atau asam format digunakan sebagai alternatif akseptor elektron oleh metanogen. Reaksi-reaksi ini sering terjadi pada arkea yang tinggal di usus. Asam asetat juga dipecah menjadi metana dan karbon dioksida secara langsung oleh arkea asetotrofik. Asetotrof adalah arkea dalam ordo Methanosarcinales dan merupakan bagian utama dari komunitas mikroorganisme yang menghasilkan biogas.[131]

Arkea lainnya menggunakan CO2 di atmosfer sebagai sumber karbon dalam proses yang disebut fiksasi karbon (mereka disebut autotrof). Proses ini melibatkan bentuk yang sangat termodifikasi dari siklus Calvin[133] atau jalur metabolik yang baru ditemukan disebut siklus 3-hidroksipropionat/4-hidroksibutirat.[134] Crenarchaeota juga menggunakan siklus Krebs terbalik, sementara Euryarchaeota juga menggunakan jalur reduktif asetil-KoA.[135] Fiksasi karbon ini didukung oleh sumber energi anorganik. Tidak ada arkea yang diketahui melakukan fotosintesis.[136] Sumber energi arkea sangat beragam dan berkisar dari oksidasi amonia oleh Nitrosopumilales[137][138] sampai oksidasi hidrogen sulfida atau unsur belerang oleh spesies Sulfolobus, yang menggunakan oksigen atau ion logam sebagai akseptor elektron.[126]
Arkea fototrofik menggunakan cahaya untuk menghasilkan energi kimia dalam bentuk ATP. Pada Halobacteria, pompa ion yang diaktifkan oleh cahaya, seperti bakteriorodopsin dan halorodopsin, menghasilkan gradien ion dengan cara memompa ion keluar dari sel melintasi membran plasma. Energi yang tersimpan dalam gradien elektrokimia kemudian diubah menjadi ATP oleh ATP sintase.[93] Proses ini merupakan bentuk fotofosforilasi. Kemampuan pompa untuk menggerakkan ion melintasi membran bergantung pada perubahan yang diberdayai cahaya dalam struktur kofaktor retinol yang ada di tengah protein.[139]
Remove ads
Genetika
Ringkasan
Perspektif
Arkea biasanya memiliki kromosom melingkar tunggal,[140] dengan ukuran hingga 5.751.492 pasangan basa pada Methanosarcina acetivorans,[141] genom arkea terbesar yang diketahui. Sepersepuluh dari ukuran ini adalah genom kecil berupa 490.885 pasangan basa pada Nanoarchaeum equitans, genom arkea terkecil yang dikenal; genom ini diperkirakan hanya berisi 537 gen penyandi-protein.[142] Potongan independen kecil dari DNA, yang disebut plasmid, juga ditemukan dalam arkea. Plasmid dapat ditransfer antarsel dengan kontak fisik, dalam proses yang mungkin mirip dengan konjugasi bakteri.[143][144]

Arkea secara genetik berbeda dari bakteri dan eukariota; sampai dengan 15% dari protein yang disandi oleh setiap genomnya merupakan protein yang unik untuk domain Archaea, meskipun sebagian besar gen yang unik tidak memiliki fungsi yang diketahui.[146] Dari sisa protein unik yang fungsinya telah diidentifikasi, sebagian besar dimiliki Euryarchaea dan terlibat dalam metanogenesis. Protein yang sama-sama dimiliki arkea, bakteri, dan eukariota membentuk sari pati fungsi sel, yang terutama berkaitan dengan transkripsi, translasi, dan metabolisme nukleotida.[147] Sifat arkea lain yang khas adalah organisasi gen yang fungsinya berhubungan—seperti enzim yang mengkatalisis langkah dalam jalur metabolik yang sama ke dalam operon baru, dan perbedaan yang besar dalam gen tRNA dan tRNA sintetase aminoasilnya.[147]
Transkripsi pada arkea lebih menyerupai transkripsi eukariota dibandingkan transkripsi bakteri, dengan RNA polimerase arkea sangat mirip dengan enzim serupa pada eukariota;[140] sementara translasi pada arkea menunjukkan tanda-tanda kesamaan, baik pada bakteri maupun eukariota.[148] Meskipun arkea hanya memiliki satu jenis RNA polimerase, struktur dan fungsinya dalam transkripsi tampaknya dekat dengan RNA polimerase II pada eukariota, dengan kumpulan protein yang sama (faktor transkripsi umum) yang mengarahkan pengikatan RNA polimerase ke promotor gen.[149] Namun, faktor transkripsi arkea lainnya lebih dekat dengan faktor transkripsi yang ditemukan pada bakteri.[150] Modifikasi setelah-transkripsi arkea lebih sederhana dibandingkan eukariota karena sebagian besar gen arkea tidak memiliki intron, meskipun ada banyak intron pada gen RNA transfer dan RNA ribosomal mereka,[151] dan intron mungkin ada pada beberapa gen penyandi-protein.[152][153]
Transfer gen dan pertukaran genetik
Halobacterium volcanii, sebuah arkea halofilik ekstrem, membentuk jembatan sitoplasma di antara sel-sel yang sepertinya akan digunakan untuk mentransfer DNA dari satu sel ke sel lain dalam dua arah.[154] Ketika arkea hipertermofilik Sulfolobus solfataricus[155] dan Sulfolobus acidocaldarius[156] terpapar radiasi sinar ultraungu (UV) yang merusak DNA atau bleomisin atau mitomisin C, agregasi seluler yang spesifik pada spesies akan diinduksi. Agregasi pada S. solfataricus tidak dapat diinduksi oleh stres fisik lainnya, seperti pH atau perubahan suhu,[155] yang menunjukkan bahwa agregasi diinduksi secara khusus oleh kerusakan DNA. Ajon dkk.[156] menunjukkan bahwa agregasi seluler yang diinduksi sinar UV memediasi pertukaran penanda kromosom dengan frekuensi tinggi pada S. acidocaldarius. Tingkat rekombinasi arkea yang diinduksi melebihi kultur yang tidak diinduksi hingga tiga lipat. Frols dkk.[155][157] dan Ajon dkk.[156] berhipotesis bahwa agregasi seluler meningkatkan transfer DNA spesifik-spesies di antara sel-sel Sulfolobus dalam rangka meningkatkan perbaikan DNA yang rusak dengan cara rekombinasi homolog. Tanggapan ini mungkin merupakan bentuk primitif dari interaksi seksual yang mirip dengan sistem transformasi bakteri yang dipelajari dengan lebih baik yang juga terkait dengan transfer DNA spesifik-spesies di antara sel-sel yang mengarah pada perbaikan rekombinasi homolog dari kerusakan DNA.[158]
Virus arkea
Arkea menjadi target dari sejumlah virus yang berbeda dari virus-virus yang menyerang bakteri dan eukariota. Virus-virus ini telah dikelompokkan dalam 15–18 keluarga berbasis DNA, tetapi beberapa spesies tetap tidak terisolasi dan menunggu klasifikasi.[159][160][161] Keluarga-keluarga ini secara informal dapat dibagi menjadi dua kelompok: virus spesifik arkea dan kosmopolitan. Virus spesifik arkea hanya menargetkan spesies arkea dan saat ini mencakup 12 keluarga virus. Banyak virus berstruktur unik yang sebelumnya tidak teridentifikasi telah ditemukan dalam kelompok ini, termasuk virus berbentuk botol, spindel, gulungan, dan tetesan.[160] Meskipun siklus reproduksi dan mekanisme genom spesies virus spesifik-arkea mungkin mirip dengan virus lain, mereka memiliki karakteristik unik yang secara khusus dikembangkan karena morfologi sel inang yang mereka infeksi.[159] Mekanisme pelepasan virus mereka berbeda dari fag lainnya. Bakteriofag umumnya menjalani jalur litik, jalur lisogenik, atau campuran keduanya (meskipun jarang).[162] Sebagian besar galur virus spesifik-arkea mempertahankan hubungan yang stabil dan agak lisogenik dengan inangnya, yang tampak sebagai infeksi kronis. Hal ini melibatkan produksi dan pelepasan virion secara bertahap dan terus-menerus tanpa membunuh sel inang.[163] Prangishyili (2013) mencatat bahwa telah dihipotesiskan bahwa fag arkea berekor berasal dari bakteriofag yang mampu menginfeksi spesies Haloarchaea. Jika hipotesisnya benar, dapat disimpulkan bahwa virus DNA berunting ganda lainnya yang menyusun sisa kelompok spesifik arkea adalah kelompok unik mereka sendiri dalam komunitas virus global. Krupovic dkk. (2018) menyatakan bahwa tingginya tingkat transfer gen horizontal, tingkat mutasi yang cepat pada genom virus, dan kurangnya urutan gen universal telah membuat para peneliti mempersepsikan jalur evolusi virus arkea sebagai jejaring. Kurangnya kesamaan antara penanda filogenetik dalam jejaring ini dan virosfer global, serta hubungan eksternal dengan elemen nonvirus, mungkin menunjukkan bahwa beberapa spesies virus spesifik-arkea berevolusi dari elemen genetik seluler nonvirus (MGE).[160]
Virus-virus ini telah dipelajari secara rinci pada kelompok termofil, khususnya ordo Sulfolobales dan Thermoproteales.[164] Dua kelompok virus DNA unting tunggal yang menginfeksi arkea telah diisolasi. Satu kelompok dicontohkan oleh Halorubrum pleomorphic virus 1 ("Pleolipoviridae") yang menginfeksi arkea halofilik[165] dan yang lainnya oleh virus aeropyrum berbentuk spiral ("Spiraviridae") yang menginfeksi inang hipertermofilik (pertumbuhan optimal pada 90–95 °C).[166] Spiraviridae dilaporkan memiliki genom ssDNA yang terbesar hingga saat ini. Pertahanan terhadap virus ini mungkin melibatkan interferensi RNA dari urutan DNA repetitif yang terkait dengan gen dari virus.[167][168]
Remove ads
Reproduksi
Ringkasan
Perspektif
Arkea bereproduksi secara aseksual dengan fisi biner atau berganda, fragmentasi, atau tunas; mitosis dan meiosis tidak terjadi, jadi jika spesies arkea memiliki lebih dari satu bentuk, semua memiliki materi genetik yang sama.[93] Pembelahan sel dikendalikan dalam siklus sel; setelah kromosom sel direplikasi dan dua kromosom anak terpisah, sel akan membelah.[169] Pada genus Sulfolobus, siklus sel memiliki karakteristik yang mirip dengan sistem bakteri dan eukariota. Kromosom bereplikasi dari beberapa titik-awal (origo replikasi) menggunakan DNA polimerase yang menyerupai enzim eukariota yang setara.[170]
Pada Euryarchaea, protein pembelahan sel FtsZ, yang membentuk cincin kontraktor di sekitar sel, dan komponen septum yang dibangun di tengah sel, mirip dengan protein serupa pada bakteri.[169] Pada cren-[171][172] dan thaumarchaea,[173] mesin pembelahan sel CDV memiliki peran serupa. Mesin ini terkait dengan mesin eukariotik ESCRT-III (yang sementara paling dikenal untuk perannya dalam pemilahan sel) yang juga terlihat memenuhi peran dalam pemisahan pada sel yang membelah, yang menunjukkan peran leluhur dalam pembelahan sel.[174]
Baik bakteri dan eukariota, tetapi tidak arkea, membentuk spora.[175] Beberapa spesies Haloarchaea menjalani peralihan fenotipik dan tumbuh sebagai beberapa jenis sel yang berbeda, termasuk struktur berdinding tebal yang tahan terhadap guncangan osmotik dan memungkinkan arkea untuk bertahan hidup dalam air pada konsentrasi garam rendah, tetapi ini bukanlah struktur reproduksi dan dapat membantu mereka mencapai habitat baru.[176]
Remove ads
Ekologi
Ringkasan
Perspektif
Habitat

Arkea hidup di berbagai habitat dan kini diakui sebagai bagian besar dalam ekosistem global,[15] yang dapat berkontribusi hingga 20% dari biomassa bumi.[177] Arkea yang pertama ditemukan adalah ekstremofil.[125] Beberapa arkea mampu bertahan pada suhu tinggi, sering kali di atas 100 °C (212 °F), seperti yang ditemukan di geyser, ventilasi hidrotermal laut-dalam, dan sumur minyak. Habitat umum lainnya termasuk habitat yang sangat dingin dan air yang sangat asin, asam, atau alkali. Selain itu, arkea juga mencakup mesofil yang tumbuh dalam kondisi ringan, seperti di tanah rawa, air limbah, samudra, saluran usus hewan, dan tanah.[15]
Arkea-arkea ekstremofil merupakan anggota dari empat kelompok fisiologis utama, yaitu halofili, termofili, alkalifili, dan asidofili.[178] Kelompok-kelompok ini tidak komprehensif atau spesifik terhadap filum, mereka juga tidak saling eksklusif karena beberapa arkea digolongkan dalam beberapa kelompok. Meskipun demikian, pengelompokan ini menjadi titik awal yang berguna untuk klasifikasi.[179]
Halofili, termasuk genus Halobacterium, hidup di lingkungan yang sangat asin seperti danau garam dan jumlahnya melebihi bakteri di lingkungan dengan salinitas yang lebih besar dari 20–25%.[125] Termofil tumbuh paling baik pada suhu di atas 45 °C (113 °F), di tempat-tempat seperti mata air panas; arkea hipertermofil tumbuh optimal pada suhu yang lebih besar dari 80 °C (176 °F).[180] Arkea Methanopyrus kandleri galur 116 bahkan dapat bereproduksi pada 122 °C (252 °F), suhu tertinggi yang tercatat dari organisme mana pun.[181]
Arkea lain hidup di kondisi yang sangat asam atau basa.[178] Sebagai contoh, salah satu arkea asidofil paling ekstrem adalah Picrophilus torridus, yang tumbuh pada pH 0, yang setara dengan berkembang di 1,2 molar asam sulfat.[182]
Ketahanan pada lingkungan yang ekstrem ini telah membuat arkea menjadi fokus spekulasi tentang kemungkinan sifat kehidupan ekstraterestrial.[183] Beberapa habitat ekstremofil tidak berbeda dengan Mars,[184] sehingga muncul kemungkinan bahwa mikrob yang sesuai bisa saja ditransfer ke antara planet-planet di meteorit.[185]
Beberapa penelitian menunjukkan bahwa arkea tidak hanya ada di lingkungan mesofilik dan termofilik tetapi juga hidup, kadang-kadang dalam jumlah yang tinggi, pada suhu rendah. Misalnya, arkea banyak ditemukan di lingkungan lautan dingin seperti laut kutub.[186] Lebih signifikan lagi adalah temuan sejumlah besar arkea di seluruh lautan di dunia pada habitat nonekstrem di antara komunitas plankton (sebagai bagian dari pikoplankton).[187] Meskipun arkea ini dapat hadir dalam jumlah yang sangat tinggi (hingga 40% dari biomassa mikrob), hampir tidak ada spesies ini yang telah diisolasi dan dipelajari dalam kultur murni.[188] Akibatnya, pemahaman tentang peran arkea di ekologi laut masih dasar sehingga pengaruh penuh mereka pada siklus biogeokimia global sebagian besar masih belum diselidiki.[189] Beberapa Crenarchaeota laut mampu melakukan nitrifikasi, yang menunjukkan bahwa organisme ini dapat memengaruhi siklus nitrogen samudra,[137] meskipun Crenarchaeota laut ini juga dapat menggunakan sumber-sumber energi lain.[190]
Sejumlah besar arkea juga ditemukan dalam sedimen yang menutupi dasar laut, dengan organisme ini membentuk sebagian besar sel hidup pada kedalaman lebih dari 1 meter di bawah dasar laut.[191][192] Telah dibuktikan bahwa di semua sedimen permukaan samudra (dari kedalaman air 1.000 sampai 10.000 m), dampak infeksi virus lebih tinggi pada arkea dibandingkan pada bakteri; sementara itu, lisis arkea yang diinduksi virus menyumbang hingga sepertiga dari total biomassa mikrob yang terbunuh, yang menghasilkan pelepasan ~0,3 hingga 0,5 gigaton karbon per tahun secara global.[193]
Peran dalam pendauran kimia
Arkea mendaur ulang unsur-unsur seperti karbon, nitrogen, dan belerang di berbagai habitat mereka.[194] Arkea terlibat dalam banyak langkah padasiklus nitrogen, yang meliputi reaksi yang dapat menghapus nitrogen dari ekosistem (seperti respirasi berbasis nitrat dan denitrifikasi) serta proses yang mendatangkan nitrogen (seperti asimilasi nitrat dan fiksasi nitrogen).[195][196] Para peneliti menemukan keterlibatan arkea pada reaksi oksidasi amonia. Reaksi-reaksi ini sangat penting di lautan.[138][197] Arkea juga tampaknya penting untuk oksidasi amonia di tanah. Mereka menghasilkan nitrit, dan mikrob lainnya mengoksidasi ke nitrat. Tumbuhan dan organisme lain mengonsumsi nitrat.[198]
Pada daur belerang, arkea yang tumbuh dengan mengoksidasi senyawa sulfur melepaskan elemen ini dari batuan agar tersedia bagi organisme lain. Namun, arkea yang melakukan hal ini, seperti Sulfolobus, menghasilkan asam sulfat sebagai produk limbah, dan pertumbuhan organisme ini di tambang yang ditinggalkan dapat berkontribusi pada air asam tambang dan kerusakan lingkungan lainnya.[199]
Dalam siklus karbon, arkea metanogen mengeliminasi hidrogen dan memainkan peran penting dalam pembusukan bahan organik dengan populasi mikroorganisme yang berperan sebagai dekomposer dalam ekosistem anaerob, seperti sedimen, rawa-rawa, dan pengolahan air limbah.[200]
Interaksi dengan organisme lain

Interaksi yang diketahui dengan baik antara arkea dan organisme lainnya bersifat mutual atau komensal. Tidak ada contoh yang jelas dan dikenal untuk arkea yang bersifat patogenik atau parasitik.[201][202] Namun, beberapa spesies metanogen diperkirakan terlibat dalam infeksi di mulut,[203][204] dan Nanoarchaeum equitans mungkin merupakan parasit bagi spesies arkea lain karena hanya bertahan dan bereproduksi dalam sel-sel Crenarchaeon Ignicoccus hospitalis,[142] dan tampaknya tidak memberikan manfaat bagi inangnya.[205] Metanogen berperan sebagai mitra utama yang mendukung proses fermentasi bakteri yang didasarkan pada sintrofi yang bekerja dalam kondisi kekurangan energi.[206]
Mutualisme
Salah satu contoh mutualisme yang dipahami dengan baik adalah interaksi antara protozoa dan arkea metanogen dalam saluran pencernaan hewan yang mencerna selulosa, seperti ruminansia dan rayap.[207] Dalam lingkungan anaerobik ini, protozoa memecah selulosa tanaman untuk memperoleh energi. Proses ini melepaskan hidrogen sebagai produk limbah, tetapi hidrogen dalam tingkat tinggi mengurangi produksi energi. Ketika metanogen mengubah hidrogen menjadi metana, protozoa diuntungkan dengan lebih banyak energi.[208]
Dalam protozoa anaerobik, seperti Plagiopyla frontata, arkea berada di dalam protozoa dan mengonsumsi hidrogen yang diproduksi di hidrogenosom mereka.[209][210] Arkea juga berasosiasi dengan organisme yang lebih besar. Sebagai contoh, arkea laut Cenarchaeum symbiosum hidup dalam (merupakan endosimbion dari) spons Axinella mexicana.[211]
Komensalisme
Arkea juga bisa menjadi komensal, yang diuntungkan dari asosiasi tanpa membantu atau merugikan organisme lainnya. Sebagai contoh, metanogen Methanobrevibacter smithii adalah arkea paling umum dalam flora manusia, yang menyusun sekitar satu dari sepuluh dari semua prokariota dalam usus manusia.[212] Pada rayap dan manusia, metanogen ini mungkin sebenarnya mutualis, berinteraksi dengan mikrob lain dalam usus untuk membantu pencernaan.[213] Dalam tubuh manusia, arkea ditemukan di usus, rongga mulut, dan kulit.[214] Komunitas arkea juga berhubungan dengan berbagai organisme lain, seperti di permukaan karang,[215] dan di wilayah tanah yang mengelilingi akar tanaman (rizosfer).[216][217]
Parasitisme
Meskipun Arkeaa secara tradisional tidak dikaitkan dengan patogenisitas, mereka sering kali memiliki genom yang memiliki kemiripan dengan patogen yang lebih umum seperti E. coli.[218] Hal ini menunjukkan adanya hubungan metabolik dan hubungan evolusioner dengan patogen kontemporer. Deteksi Arkea yang terputus-putus dalam penelitian klinis dapat dikaitkan dengan tidak adanya klasifikasi spesies yang lebih tepat untuk Arkea.[219]
Remove ads
Peran dalam teknologi dan industri
Ringkasan
Perspektif
Arkea ekstremofili, terutama yang tahan terhadap panas atau keasaman dan alkalinitas ekstrem, merupakan sumber enzim yang berfungsi dalam kondisi yang ekstrem.[220][221] Enzim-enzim ini memiliki banyak kegunaan. Misalnya, DNA polimerase termostabil, seperti Pfu DNA polimerase dari Pyrococcus furiosus, merevolusi biologi molekuler dengan memungkinkan reaksi berantai polimerase digunakan dalam penelitian sebagai teknik sederhana dan cepat untuk kloning DNA. Dalam industri, amilase, galaktosidase, dan pullulanase dari spesies Pyrococcus yang lain yang berfungsi pada suhu di atas 100 °C (212 °F) sehingga memungkinkan pengolahan makanan pada suhu tinggi, seperti produksi susu rendah laktosa dan whey.[222] Enzim dari arkea termofilik ini juga cenderung sangat stabil dalam pelarut organik, sehingga dapat digunakan dalam proses ramah lingkungan dalam kimia hijau yang menyintesis senyawa organik.[221] Stabilitas ini membuatnya lebih mudah untuk digunakan dalam biologi struktural. Akibatnya, enzim dari arkea ekstremofil yang ekuivalen dengan enzim bakteri atau eukariotik sering digunakan dalam studi struktural.[223]
Berbeda dengan berbagai aplikasi enzim arkea, penggunaan organisme-organisme arkea sendiri di bidang bioteknologi kurang dikembangkan. Arkea metanogen adalah bagian penting dari pengolahan limbah karena mereka adalah bagian dari komunitas mikroorganisme yang melaksanakan pencernaan anaerobik dan menghasilkan biogas.[224] Dalam pengolahan mineral, arkea asidofilik dipandang menjanjikan untuk ekstraksi logam dari bijih, termasuk emas, kobalt dan tembaga.[225]
Arkea menghasilkan antibiotik kelas baru yang berpotensi dapat digunakan. Beberapa antibiotik asal arkeosin telah dikarakterisasi, tetapi ratusan calon antibiotik lainnya diyakini keberadaannya, terutama dari Haloarchaea dan Sulfolobus. Senyawa-senyawa ini memiliki struktur yang berbeda dari antibiotik asal bakteri sehingga mereka mungkin memiliki cara kerja yang baru. Selain itu, mereka dapat memungkinkan pembuatan penanda yang dapat diseleksi yang baru untuk digunakan dalam biologi molekuler arkea.[226]
Remove ads
Lihat pula
- Produksi metana aerobik
- Bentuk kehidupan terawal yang diketahui
- Daftar genus Arkea
- Daftar genom arkea yang sudah diurutkan
Referensi
Bacaan lebih lanjut
Pranala luar
Wikiwand - on
Seamless Wikipedia browsing. On steroids.
Remove ads