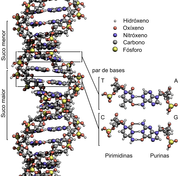
As desoxirribonucleases (DNases) son un grupo de glicoproteínas endonucleases que son encimas que catalizan a rotura por hidrólise de enlaces fosfodiéster nunha cadea de ADN, o que produce a degradación do ADN. O papel dos encimas DNases nas células inclúe a degradación do ADN extracelular (ADNec) excretado pola apoptose, a necrose e as trampas extracelulares de neutrófilos das células para axudar a reducir as respostas inflamatorias que doutro modo se desencadearían. Coñécese unha ampla variedade de desoxirribonucleases que se clasifican en dúas familias (DNase I e DNase II), que se diferencian nas súas especificidades de substrato, mecanismos químicos e funcións biolóxicas. Entre as aplicacións de laboratorio das DNases están a purificación de proteínas cando se extraen de organismos procariotas. Ademais, as DNases utilizáronse como tratamento de doenzas causadas por ADN extracelular no plasma sanguíneo. Neste campo empezaron a utilizarse tamén ensaios de DNase.
Tipos
Os dous grandes tipos de DNases que se encontran en metazoos son os das familias da desoxirribonuclease I (DNase I) e da desoxirribonuclease II (DNase II). Estas dúas familias comprenden varios subtipos.
Familia da DNase I: DNase I, DNase1L1, DNase 1L2, DNase1L3
O primeiro conxunto de DNases é o da familia da DNase I. Esta familia consta da DNase I, DNase1L1, DNase 1L2 e DNase1L3. A DNase I corta o ADN para formar dous produtos finais de oligonucleótidos con extremos 5’-fosfato e 3’-hidroxi e prodúcese principalmente en órganos do aparato dixestivo. A familia da DNase I require os catións Ca2+ e Mg2+ como activadores e exprésase selectivamente.[1] A familia da DNAse I é activa a un pH entre 6,5 e 8, normal nas células.
Familia da DNase II: DNase II ɑ e DNase II ꞵ
O segundo conxunto de DNAases é o da familia da DNase II. Esta familia consta da DNase II ɑ e a DNase II ꞵ. Igual que as DNAases I, as DNAases II cortan o ADN para formar produtos finais oligonucleótidos con extremos 5'-P e 3'-OH. Este tipo de DNAase exprésase máis abundantemente en tecidos debido á alta expresión en macrófagos pero cunha expresión limitada ao tipo celular. A diferenza da DNAase I, non necesita catións Ca2+ e Mg2+ como activadores.[2] Os encimas desta familia funcionan a pH ácido. O padrón de clivaxe das DNases II altérase en presenza de dimetil sulfóxido (DMSO), que afecta significativamente a estrutura do ADN.
Estrutura
Aínda que ambos os tipos de DNases son glicoproteínas endonucleases, a DNase I ten unha estrutura monómera tipo sándwich cunha cadea lateral de carbohidratos, mentres que a DNase II ten unha estrutura cuaternaria dímera.

Estrutura da DNase I: A DNase I é unha glicoproteína cun peso molecular de 30.000 Da e unha cadea de carbohidratos de 8-10 residuos unida á Asn18 (en laranxa na figura).[3] É unha proteína 𝛼,𝛽 con dúas follas 𝛽 de 6 febras que forman o núcleo da súa estrutura.[4] Estas dúas láminas beta da parte central corren paralelamente, e todas as outras corren antiparalelamente. As follas 𝛽 están na parte central da estrutura, mentres que as hélices 𝛼 son as colas periféricas. A DNase I contén catro petos para a unión de ións e necesita Ca2+ e Mg2+ para hidrolizar o ADN bicatenario.[5] Dous dos sitios únense fortemente ao Ca2+ mentres que os outros dous coordinan o Mg2+. Publicouse pouco sobre o número e localización dos sitios de unión para o Mg2+, aínda que se propuxo que o Mg2+ está localizado preto do peto catalítico e contribúe á hidrólise.[6] Os dous Ca2+ móstranse en vermello na imaxe. Únense á DNase I en condicións de cristalización e son importantes para a integridade estrutural da molécula ao estabilizaren o bucle superficial de Asp198 a Thr204 (en cor ciano), e limitando a rexión de alta mobilidade térmica no bucle flexible aos residuos Gly97 a Gly102 (en amarelo).

Estrutura da DNase II: A DNase II ten unha estrutura cuaternaria homodímera que pode unirse ao ADN bicatenario cunha arquitectura de abrazadeira con forma de U. O interior da abrazadeira con forma de U é bastante electropositiva, con capacidade de unirse ao ADN cargado negativamente. De xeito similar á DNase I, a estrutura da DNase II consta dunha estrutura secundaria mixta 𝛼/𝛽 con 9 hélices 𝛼 e 20 follas 𝛽.[7] A diferenza da DNase I, a DNase II non necesita ións metálicos divalentes para a catálise.[7] A estrutura consta dun protómero A (ciano) e un protómero B (verde). Cada estrutura consta de dous motivos catalíticos, que están etiquetados só no protómero B para simplificar: His100 e Lys102 compoñen o primeiro motivo (azul) e His279 e Lys281 compoñen o segundo motivo catalítico (vermello).

Mecanismo
Algunhas DNases cortan só residuos situados nos extremos das moléculas de ADN. Este tipo de exonuclease denomínase exodesoxirribonucleases. Outras cortan en calquera punto do interior da cadea e denomínanse endodesoxirribonucleases (un grupo de endonucleases.)[8] Algunhas DNases son bastante indiscriminadas en canto á secuencia de ADN na cal cortan, mentres que outras, como os encimas de restrición, son moi específicos de secuencia. Outras DNases cortan só ADN bicatenario, outros son específicos de moléculas monocatenarias e outras actúan sobre ambas.
A acción dunha DNase occorre en tres fases. A fase inicial produce múltiples amosegas na cadea de enlaces fosfodiéster. A segunda fase produce nucleótidos solubles en ácido. A terceira fase, que é a fase terminal, consiste na redución de oligonucleótidos, causando un cambio hipercrómico nos datos UV.[9]
Mecanismo da DNase I
A DNase I ten como diana predominante o ADN bicatenario e, en moita menor medida, algúns ADN monocatenarios. A DNase I cataliza o corte do ADN non específico rompendo enlaces fosfodiéster nunha das febras. O seu sitio de corte sitúase entre o átomo de oxíxeno 3′ e o átomo de fósforo adxacente, rendendo oligonucleótidos 3′-hidroxilo e 5′-fosforilo con inversión da configuración no fósforo. O encima DNase depende da presenza dun catión divalente, que adoita ser o Ca2+, para funcionar correctamente. O sitio activo da DNase I comprende dous residuos de histidina (His134 e His252) e outros dous residuos ácidos (Glu78 e Asp212), todos os cales son esenciais para a catálise ácido-básica dos enlaces fosfodiéster.[10]
Mecanismo da DNase II
A desoxirribonuclease II (DNase II) tamén se coñece como desoxirribonuclease ácida porque ten unha actividade óptima no ambiente de pH baixo (ácido) dos lisosomas, onde é típico encontrala nos eucariotas superiores. Algunhas formas de DNase II recombinante mostran un alto nivel de actividade a pH baixo en ausencia de ións metálicos divalentes, similar á da DNase II eucariota.[7] A diferenza da DNase I, a DNase II cliva o enlace fosfodiéster entre o átomo de oxíxeno 5' e o átomo de fósforo adxacente, rendendo nucleótidos 3΄-fosforilados e 5΄-hidroxilo.
Aplicacións
Aplicacións de laboratorio
A DNase utilízase comunmente cando se purifican proteínas extraídas de organismos procariotas. A extracción de proteínas a miúdo require a degradación da membrana celular. É común lisar a fráxil membrana, liberando o ADN, que non se desexa neste proceso, xunto coas proteínas desexadas. O extracto resultante de ADN e proteínas é moi viscoso e difícil de purificar, para o cal se engade DNase para degradar o ADN.[11] Así, o ADN é hidrolizado pero as proteínas non son afectadas e o extracto pode seguir coas seguintes fases da purificación.
Tratamento
O ADN extracelular (ADNec ou ecDNA) é o ADN que se encontra na circulación sanguínea. Aparece como resultado da apoptose, necrose ou trampas extracelulares de neutófilos que afectan a células do sangue e de tecidos, pero pode tamén orixinarse pola secreción activa de células vivas. O ADNec e as súas proteínas que se unen ao ADN poden activar os receptores que detectan o ADN, os receptores de recoñecemento de padróns (PRRs). Os PRRs poden estimular vías que causan unha resposta inflamatoria inmune. Como resultado, varios estudos de doenzas inflamatorias atoparon que hai altas concentracións de ADNec no plasma sanguíneo. Por esta razón, a DNase é un posible tratamento para a redución da ADNec no plasma sanguíneo. As DNases poden ser excretadas tanto intracelularmente coma extracelularmente e poden cortar os enlaces fosfodiéster do ADN. Esta función pode usarse para manter unha baixa concentración de ADNec, tratando así a inflamación. As enfermidades que son o resultado da presenza deste residuo de ADN no sangue foron tratadas usando as "propiedades degradantes" da DNase. Os estudos realizados mostraron que a DNase pode actuar como un tratamento facendo diminuír a viscosidade do moco.[12][13] A administración de DNase varía dependendo da doenza. Foi administrado oralmente, intrapleuralmente, por vía intravenosa e intraperitoneal e por inhalación.[14] Varios estudos continúan examinando a aplicación da DNase como tratamento, así como unha forma de monitorizar a saúde. Por exemplo, recentemente a DNase derivada de bacterias patóxenas utilizouse como indicador para monitorizar as infeccións de feridas.[15]
Doenzas respiratorias
A fibrose quística é un trastorno xenético que afecta a produción de moco, suor e fluídos dixestivos, causando que sexan máis viscosos en vez de lubricantes. Os encimas DNases poden ser inhalados usando un nebulizador por pacientes de fibrose quística. Son útiles porque os glóbulos brancos se acumulan no moco e, cando se destrúen, liberan ADN, o cal aumenta o carácter pegañento do moco. Os encimas DNases degadan ese ADN e así o moco é moito máis fácil de eliminar dos pulmóns. Concretamente, a DNase I, tamén coñecida como o fármaco aprobado pola FDA estadounidense Pulmozyme (tamén coñecido como dornase alfa), utilízase como tratamento para incrementar a función pulmonar.
Outras doenzas respiratorias como a asma,[16] empiema pleural,[12] e enfermidade pulmonar obstrutiva crónica (EPOC) tamén se ven positivamente afectadas polas propiedades das DNases.
Ademais, estudos recentes mostran que o activador do plasminóxeno tisular (tPA) intrapleural, unha proteína que é responsable da disolución de coágulos sanguíneos, combinada coa desoxirribonuclease incrementa a drenaxe pleural, diminúe a estancia hospitalaria e a necesidade de cirurxía nas efusións parapneumónicas e no empiema.
Outras doenzas
A sepse é unha doenza inflamatoria que pode ser mortal causada pola resposta extrema do corpo a unha infección. O corpo empeza atacándose a si mesmo como unha resposta inflamatoria que afecta a todo o corpo. Como resultado, aparecen altos niveis de ADNec na corrente sanguínea e, por tanto, os investigadores pensaron na DNase como tratamento apropiado. Os estudos mostraron que a DNase alteraba as trampas extracelulares de neutrófilos e diminuía as respostas inflamatorias. Cómpre facer máis investigacións sobre o tipo e tempo de administración para establecer a DNase como un tratamento oficial.[17][18][19]
O lupus eritematoso sistémico é unha enfermidade autoinmune que ten como resultado a xeración de autoanticorpos causando unha inflamación que causa danos nos órganos, articulacións e especialmente nos riles. O lupus eritematoso sistémico está ligado a baixos niveis de DNase I a medida que células apoptóticas se converten en autoantíxenos nesta doenza. A DNase I investigouse como un posible tratamento para diminuír a cantidade de refugallos apoptóticos no corpo humano. Os estudos sobre este tratamento obtiveron resultados contraditorios e están realizándose novas investigacións sobre os beneficios terapéuticos da DNase I.[14][18][20]
Tratamento antitumoral. A DNase ten efectos antitumorais debido á súa capacidade de degradar o ADN. No sangue de pacientes de cancro atópanse altos niveis de ADN, o que suxire que a DNase I podería ser un posible tratamento. Aínda se comprende mal por que hai altos niveis de ADNec e se a DNase pode actuar ou non como tratamento efectivo. Varios estudos en ratos mostraron resultados positivos na progresión antitumoral utilizando DNase I intravenosa. Porén, haberá que facer máis investigacións antes de introducilo como tratamento.[21][14]
Ensaios
O ADN absorbe a luz ultravioleta (UV) cunha absorbancia máxima na lonxitude de onda preto de 260 nm. Esta absorción débese aos electróns pi nas bases aromáticas do ADN. No ADN bicatenario ou mesmo en rexións do ARN onde hai unha estrutura de dobre cadea, as bases están empilladas paralelamente unhas a outras e o solapamento dos orbitais moleculares das bases produce unha diminución da absorbancia de luz UV. Ese fenómeno chámase efecto hipercrómico. Cando a DNase libera nucleótidos do ADN bicatenario, as bases xa non están empilladas como o estaban no ADN, así que o solapamento de orbitais minimízase e a absorbancia UV aumenta. Este incremento de absorbancia é no que se basea a unidade de Kunitz da actividade da DNase. Unha unidade de Kunitz defínese como a cantidade de encima engadido a 1 mg/ml de ADN de esperma de salmón que causa un incremento de absorbancia de 0,001 por minuto na lonxitude de onda de 260 nm cando actúa sobre ADN moi polimerizado a 25 °C en tampón 0,1 M NaOAc (pH 5,0). O nome da unidade é un recoñecemento ao bioquímico ruso-americano Moses Kunitz, que propuxo o test estándar en 1946.[22]
Unha preparación encimática estándar debería correr en paralelo cunha estandarización descoñecida porque a estandarización de preparacións do ADN e o seu grao de polimerización en solución non é posible.
Difusión de encima radial sinxela (SRED, Single Radial Enzyme Diffusion) Este simple método para a medida da actividade da DNase I foi introducida por Nadano et al. e está baseado na dixestión do ADN en xel de agarosa pola DNase, que está presente en mostras picadas no xel.[14] A actividade de DNase está representada polo tamaño dun pozo circular nunha capa de xel de agarosa, no cal o ADN tinguido polo bromuro de etidio está distribuída uniformemente. Despois da incubación fórmase unha zona escura circular a medida que o encima difunde polo xel desde o pozo radialmente e corta o ADN. O SRED experimentou moitas modificacións, que levan ao incremento na sensibilidade e seguridade, como a substitución do bromuro de etidio por SYBR Green I ou outra tinguidura para xel de ADN.[23]
Ensaio da actividade colorimétrica da DNase I
O ensaio de actividade de ADNase I colorimétrica realízase para a estimación da estabilidade da DNase I recombinante humana (Pulmozyme). O método foi axustado a partir dun ensaio de actividade encimática final colorimétrica baseado na degadación dun complexo ADN/verde de metilo.[24]