在大自然中,我们可以观察到各种各样不同的图案、形式或规律,它们可被统称为“自然界中的模式”。这些“模式”会在不同的现实情况中重复出现,当中有部分更可以通过运用数学建模去理解。自然的模式会以例如对称、树状、螺旋、曲折、波状、泡沫、镶嵌、裂纹和班纹等等不同方式呈现。[1] 在历史上,早期的希腊哲学家如柏拉图、毕达哥拉斯和恩培多克勒等,已开始研究不同种类的模式;他们都试图去解释自然界中隐藏着的秩序。而随着时间的推移,现代人亦逐渐发展并加深了对各种自然模式的理解。

.jpg)
例如在19世纪时,比利时物理学家约瑟夫·普拉托对肥皂膜作出了研究,并得出了“极小曲面”的概念。德国生物学家和艺术家恩斯特·海克尔又绘画了数百种海洋生物,以展现它们的对称之美。苏格兰生物学家达西·汤普森更带头研究植物和动物的生长模式,利用了简单的数学方程式去解释螺旋形态的生长。到20世纪,英国数学家艾伦·图灵预测了产生斑点和条纹图案(Turing pattern)的“形态发生”机制。匈牙利生物学家阿里斯蒂德·林登马耶(Aristid Lindenmayer)和法裔美国数学家本华·曼德博则展示了“分形”数学(fractals)如何可以模拟出植物的生长模式。
总的来说,我们可以利用科学中的数学、物理学和化学知识,去理解自然界中不同层次和尺度的模式。又可以通过生物学上的自然选择和性选择过程,来解释各种生物在形体上呈现的独特模式。在有关“模式形成”(pattern formation)的研究当中,科学家更是利用了计算机模型,去广泛地模拟和探索各种各样模式背后的原理。
历史
早期的希腊哲学家试图解释自然界中的秩序,造就了现代人关于“自然模式”的概念。例如,毕达哥拉斯(c.570 – c.495 BC)提出自然界中的模式(例如音乐的和声)是由数字产生的,他认为数字是“存有”的基本组成部分。[a] 恩培多克勒(c.494 – c.434 BC)在一定程度上预视了达尔文对生物结构进化的解释。[b] 柏拉图(c.427 – c.347 BC) 又论述了在自然中“共相”(universals)的存在;他认为这些共相由“理想形式”(eidos)组成,而具体事物只不过是不完美的复制品(举例说,一朵花可能大致上是圆形的,但它绝不是一个完美的圆形)。[2] 泰奥弗拉斯托斯(c.372 – c.287 BC) 则指出,“具有平叶的植物,其叶子均有规则地排列”;而老普林尼(23 – 79 AD)更看出叶子的图案是随着主干成圆形排列的。[3] 多个世纪后,列奥纳多·达·芬奇(1452 – 1519)注意到树叶图案的螺旋形排列,以及树干会随着年龄的增长显示出年轮,并提出了一个法则,可以用来了解树枝横截面的总面积。[3][4]
1202 年,列奥纳多·斐波那契通过他的著作《计算之书》(Liber Abaci),首次将“斐波那契数列”介绍给西方世界。[5] 在书中,斐波那契提出了一个关于“理想化”兔子种群增长的思想实验。[6]之后, 约翰内斯·开普勒(1571–1630)指出自然界的模式当中存在着斐波那契数列,并用它来解释了一些花卉的五边形形状。[3] 到1658 年,英国医生和哲学家托马斯·布朗爵士在《居鲁士花园》(The Garden of Cyrus)中讨论了“自然怎样几何化”,引用了涉及数字“5”的毕达哥拉斯命理学,和梅花形图案当中的柏拉图理形(论文的中心章节叙述了植物学中有关梅花形的例子和观察)。[7] 1754 年,查尔斯·邦尼特观察到植物的螺旋形叶序经常以顺时针或逆时针方向的黄金比例序列呈现。[3] 卡尔·弗里德里希·辛柏(Karl Friedrich Schimper)和他的朋友亚历山大·布劳恩(Alexander Braun)在 1830 年的工作中,开展了对于叶序的数学研究;奥古斯特·布拉维和他的兄弟路易在 1837 年将叶序比率与斐波那契数列联系起来,还注意到它出现在松果和凤梨的图案之中。[3] 德国心理学家阿道夫·蔡辛(Adolf Zeising)在他 1854 年出版的书中,更详细地探讨了在植物中各部位的排列、动物的骨骼、它们的静脉和神经的分支模式、以及晶体等等的例子当中,所表现出的黄金比例。[8][9][10]
在 19 世纪,比利时物理学家约瑟夫·普拉托(1801 – 1883 年)提出了“给定边界是否都具有极小曲面”这个数学问题,现在该问题以他的名字命名。他深入地研究了肥皂薄膜的特性,更制定了普拉托定律,去描述泡沫中的薄膜所形成的结构。[11] 开尔文勋爵在 1887 年找到了将等体积的胞体装叠为泡沫结构的最有效方法;他的解决方案只使用了一种立体,即具有非常轻微弯曲面的截角八面体堆砌,以满足普拉托定律。直到 1993 年,当丹尼斯·韦尔(Denis Weaire)和罗伯特·费伦(Robert Phelan)提出韦尔—费伦结构时,才找到更好的解决方案(为2008年夏季奥运会建造的北京国家游泳中心的外墙设计便是应用了该种结构)。[12] 另外,恩斯特·海克尔(1834 – 1919) 绘制了关于海洋生物的精美插图(特别是放射虫),凸显了它们的对称性,以支持他的“仿达尔文”进化论。[13] 美国摄影师威尔逊·本特利(Wilson Bentley) 则于 1885 年拍摄了第一张有关雪花的显微照片,展现了微观世界中美丽的自然模式。[14]
到 20 世纪,阿图·哈里·查尔赫(A. H. Church)在他 1904 年的书中继续研究了叶序的模式。[15] 1917 年,达西·汤普森发表了《论生长与形态》(On Growth and Form);他对叶序和斐波那契数列的阐述(即是植物螺旋生长形式的数学关系),表明了简单的方程式足以描述动物角和软体动物壳的螺旋生长模式。[16]到 1952 年,计算机科学家艾伦·图灵(1912 – 1954)撰写了《形态发生的化学基础》(The Chemical Basis of Morphogenesis),分析了在称为形态发生的过程中,在活生物体中产生出模式所需的机制。[17] 他预测了化学振荡反应现象,特别是别洛乌索夫-扎博廷斯基反应。图灵认为,这些“激活剂-抑制剂”机制可以在动物身上产生出条纹和斑点的图案(称为“图灵图案”),亦可以促成在植物叶序中看到的螺旋形结构模式。[18] 1968 年,匈牙利理论生物学家阿里斯蒂德·林登马耶(Aristid Lindenmayer)(1925 – 1989)更开发了“L-系统”,这是数学上的一种形式文法,可用于以分形的方式对植物的生长模式进行数学建模。[19] L-系统有一个符号字母表,可以使用“产生式规则”组合以构建更大的字符串,同时亦有机制将生成的字符串转换为几何结构。到 1975 年,基于戈特弗里德·莱布尼茨、格奥尔格·康托尔、海里格·冯·科赫和瓦茨瓦夫·谢尔宾斯基等人在数个世纪以来在“模式数学”上的缓步发展,本华·曼德博写出了一篇著名的论文《英国的海岸线有多长?统计自相似和分数维度》,将该类数学思想结晶成“分形”的概念。[20]

.jpg)

模式的成因

在诸如兰花、蜂鸟和孔雀尾巴等生物身上可见到的丰富“设计”,具有艺术家难以仿效的形状、图案和色彩之美。[21] 人们在自然界中所感知到的美,背后有着不同层次的科学原因,其中主要是令模式得以实现的的数学原理,以及在物种当中令模式得以演化的自然选择原理。[22]
数学旨在发现和解释各种各样抽象的模式或规律。[23][24] 很多自然界中可见的模式可以在混沌理论、分形、对数螺线、拓扑学和其他数学范畴中找到解释。例如,利用L-系统可建构出可信的数学模型,去模拟不同种类树木的生长模式。[19]
物理学定律将数学上的抽象模式应用到现实世界中,而暂时将现实当作是完美的、或理想化的。例如,当晶体没有位错等结构缺陷并且完全对称时,它就是完美的晶体。但精确的数学上的完美只能近似真实的事物。[25] 而实际上,自然界中可见的模式均受物理定律所支配;例如,可以利用流体动力学来解释曲流的形成过程。
在生物学中,自然选择可以令生物发展出各种模式;而当中有着不同的原因,包括伪装[26]、性择[26]、和不同类型的传信行为,例如拟态[27]和清除共生[28]。在植物中,百合等虫媒花的形状、颜色和图案,进化至可以易于吸引蜜蜂等昆虫;例如一些呈放射状的颜色和条纹图案(有些只在紫外光下才能看到),成为在远处也可以看到的蜜源标记。[29]
模式的类型
对称性在生物中无处不在。动物主要是左右对称或镜像对称的,植物的叶子和某些花(例如兰花)也是如此。[30] 植物通常具有径向或旋转对称性,许多花和一些动物群(如海葵)也是如此。[30] 有些棘皮动物具有五重对称性,包括海星、海胆和海百合。[31]
在非生物中,雪花具有突出的六重对称性;每个薄片的结构记录了其结晶过程中所遇到的环境变化,其六个臂中每个的生长模式几乎相同。[32] 晶体一般具有多种对称性和晶体习性;它们可以是立方体或八面体,但真正的晶体不能具有五重对称性(与准晶体不同)。[33] 在非生物中亦可见到不同尺度的旋转对称性,包括当水滴落入池塘时形成的冠形飞溅图案,[34] 以及像土星这样的行星的球形和星环。[35]
生物上的对称有多种成因。径向对称适合像海葵这种在成虫阶段不会移动的生物,因为食物和威胁可能来自任何方向。但向一个方向移动的动物必然有上和下侧、头和尾,因此也有左和右侧。头部会出现特化,有嘴和感觉器官(头颅化),而身体变得左右双侧对称(尽管体内器官不必如此)。[36] 而颇令人费解的是棘皮动物具有五重(五辐射)对称性的原因。早期的棘皮动物是双侧对称的,它们的幼虫仍然如此。Sumrall 和 Wray 认为,旧对称性的丧失既有生物发展上的原因,也有生态环境上的原因。[37]

_cropped.jpg)







达芬奇在意大利文艺复兴时期描述了树木的分枝模式。他在《绘画论》中说:
一棵树在其每个阶段高度的所有树枝的切面宽度算在一起时,与[在它们下面的]树干的切面宽度相等。[38]
更普遍的说法是,当母枝分岔成两个或多个子枝时,子枝的切面面积加起来就是母枝的切面面积。[39] 这等于说,如果一个母枝分裂成两个子枝,则母枝和两个子枝的横截面直径的长度值可以构成一个直角三角形。一种解释是,这个生长模式可以让树木更好地抵御强风。[39] 生物力学模型的模拟结果与这个规则相符。[40]
分形是无限自相似的、具有分形维数的迭代数学结构。[20][41][42] 无限迭代在自然界中是不可能的,因此现实中所有的“分形”模式都只是近似的。例如,蕨类植物和伞形科植物的叶子只有2、3 或 4 层的自相似(羽状)模式。蕨类植物的生长模式可以出现在植物和动物身上,例如苔藓虫、珊瑚、水螅纲(如 Sertularia argentea)、以及一些非生物事物,例如放电现象。L-系统分形(数学模型)可以通过改变少量参数来模拟不同种类的树木生长模式,这些参数包括分支角度、节点或分支点之间的距离(节间长度)、以及每个分支点的分支数量。[19]
分形模式在自然界中广泛存在,出现在云、河网、地质断层线、山脉、海岸线[43]、动物体色、雪花[44]、晶体[45]、血管分支[46]、浦肯野细胞[47]、肌动蛋白细胞骨架[48]、和海浪[49]等等,多种多样的现象中。


.jpg)
.jpg)



螺旋在植物和一些动物中很常见,尤其是软体动物。例如在鹦鹉螺(一种头足类软体动物)中,其壳的每个腔室都是下一个腔室的近似副本,按固定比例缩放并以对数螺旋形排列。[50] 基于现代对分形的理解,生长上呈现的螺旋状可以被视为自相似模式的一种特例。[51]
植物中的螺旋模式可以在各种生长情况中看到,例如叶序(茎叶的排列),和其他部分的排列(斜列线[52]),如向日葵等菊科植物的花头和种子头,或菠萝[15][53]:337和蛇皮果等水果的形态,以及松果中的鳞片模式(其中多个螺旋沿顺时针和逆时针运行)。这些模式在不同的科学知识层面上都有解释——数学、物理、化学、生物学——每一个层面都各自正确,而合起来看时全都是必要的。[54] 叶序中的螺旋模式可以从斐波那契比率生成:斐波那契数列为 1、1、2、3、5、8、13...(每个后续数字是前两个数字的和)。举例说,当叶子在茎上交替排列时,如果螺旋每旋转一圈会接触到两片叶子,这模式或比例就称为 1/2。在榛子中,比例是 1/3;在杏子中是 2/5;梨是 3/8;杏仁则是 5/13。[55]
在向日葵和雏菊的圆盘形花序中,小花沿费马螺线排列,但这只是看似如此,因为连续的小花其实间隔很远,分隔角度为黄金角即 137.508°(以黄金比例划分圆的 360°);当花头成熟、所有小花都长到相同大小时,这个特别的花头间距便会组成更明显的、符合斐波那契数字的螺旋图案。[56]
从物理学的角度来看,螺旋是能量最低的形态[57],它通过动态系统中的自组织过程自发出现。[58] 从化学的角度来看,螺旋模式可以通过反应-扩散过程产生,当中包括活化和抑制作用。叶序受到不同的化学机制所控制,包括一些可调节植物激素“生长素”浓度的蛋白质(生长素会激活分生组织生长),以及一些可调控茎周围芽的相对角度的其他机制。[59] 而从生物学的角度来看,在任何给定的空间中尽可能远地布置叶子是自然选择的结果,因为这样可以让植物最大程度地获取资源,尤其是用于光合作用的阳光。[53]

.jpg)


.JPG)

_(4308584755).jpg)
.jpg)
在数学上,一个动力系统如果对初始条件高度敏感(所谓的“蝴蝶效应”[60]),则它便是所谓混沌的,这需要符合拓扑混合和稠密周期轨迹的数学特性。[61]
基本上,除了分形,混沌理论亦普遍地影响着自然界中的模式。混沌与分形之间存在着关系——数学上,混沌系统中的奇异吸引子具有分形维数。[62] 一些元胞自动机(即可以生成模式的一些简单数学规则集)表现出混沌的行为,尤其是史蒂芬·沃尔夫勒姆所提出的“规则30”。[63]
涡街是由流体(最常见的是空气或水)流动时,在阻碍物体上的不稳定分离而产生的旋转涡流的锯齿形图案。[64] 当障碍物的大小或流体的流动速度与其粘度相比变得足够大时,平滑的流动(层流)便开始被破坏,而产生出涡街的现象。
曲流是当河流或其他渠道中的流体(通常是水)不断流过一些弯位时,所逐渐形成的蜿蜒弯道。当流动路径稍微有弯曲时,随着螺旋流将沙子和砾石等物质拖过河流到弯道内侧,每个弯道的大小和曲率就会因而增加。而弯道的外侧变得水流清彻且不受保护,因此侵蚀加速,于是在强大的正反馈作用中进一步增加了河道的蜿蜒曲折。[65]
-

-
 流动:胡安·费尔南德斯群岛(Juan Fernandez Islands)附近的云涡街
流动:胡安·费尔南德斯群岛(Juan Fernandez Islands)附近的云涡街 -

-
 蜿蜒曲折:古巴考托河的蜿蜒路径
蜿蜒曲折:古巴考托河的蜿蜒路径 -
 蜿蜒曲折:蛇的蜿蜒状的爬行
蜿蜒曲折:蛇的蜿蜒状的爬行 -
 蜿蜒曲折:体形对称的脑珊瑚(Diploria strigosa)上的图案
蜿蜒曲折:体形对称的脑珊瑚(Diploria strigosa)上的图案
.JPG)



.jpg)
_closeup.jpg)
波浪是在移动时携带着能量的扰动。机械波通过介质(空气或水)传播,使后者在波经过时发生振荡。[66] 风波是海面波,它会引発任何大型水体都一般会出现的混沌模式,尽管它们在统计上的行为可以通过风波模型来预测。[67] 当水或风中的波浪经过沙子表面时,它们会产生涟漪状的图案。当风吹过大片沙地时,地面会形成沙丘,有时更会像塔克拉玛干沙漠那样成为一片广阔的沙丘地带。沙丘可以呈现出不同的图案,包括新月形、很长的线形、星星形、圆顶形、抛物线形、和纵向(或称为“剑丘”)的形状。[68]
新月形沙丘是由风作用在沙漠地面而产生的;新月的两个角和滑波面指向顺风方向。沙子从逆风面(与水平面约成 15 度角)吹过,落到滑波面那边,在那里积聚到沙子成休止角(水平面约 35 度角)。当滑波面超过休止角时,沙子便会发生像雪崩一样的崩塌,这是一种非线性行为:持续添加少量沙子不会发生任何事情,但是在临界点再添上少量沙子却会突然引发大型崩塌。[69] 不过,在这种非线性行为之外,新月形沙丘的整体行为则更像是孤波。[70]




肥皂泡会形成一个球面,一个面积最小的表面(极小曲面)——可以围起某一体积的最小表面面积。两个气泡一起形成的形状则更复杂:两个气泡的外表面都是球形的;而这两个表面由第三个球形表面连接起来,其中较小的气泡会稍微凸出到较大的气泡之中。[11]
泡沫是一团气泡;自然界中存在着由不同物质形成的泡沫。由肥皂膜组成的泡沫遵循普拉托定律,即要求三个肥皂膜在每个边缘以 120° 相交,四个肥皂膜边缘在每个顶点以大约 109.5° 的四面体角相交。普拉托定律进一步要求薄膜是平滑和连续的,并且在每个点处具有恒定的平均曲率。例如,薄膜可以通过在一个方向(例如,从左到右)向上弯曲、同时在另一个方向(例如,从前到后)向下弯曲,而整体上保持几乎平坦。[71][72] 在现实中,具有最小表面的结构可以作为例如帐篷之用。
在活细胞的尺度上,泡沫型态很常见;例如放射虫、海绵骨针、硅鞭毛藻的外骨骼、和一种海胆(Cidaris rugosa)的方解石骨架,都像是普拉托泡沫边界的矿物铸型。[73][74] 放射虫 Aulonia hexagona 的骨架是被恩斯特·海克尔绘制过的一种美丽的海洋生物,它看起来像是一个完全由六边形组成的球体,但这在数学上是不可能的。欧拉示性数表明,对于任何凸多面体,面数加上顶点(角)数等于边数加二。这个公式的一个结论是任何封闭的六边形多面体都必须包含正好 12 个五边形,如足球、巴克敏斯特·富勒的测地穹顶或富勒烯分子。要明白这个,可以设想六边形网格就像一张铁丝网一样平坦,但是添加的每个五边形都会迫使网格弯曲(角落更少,因此网格被拉入)。[75]







镶嵌或密铺是通过在整个平面上用重复的形状铺砌而形成的图案或模式。在数学上,只有 17 种独立的方式可以密铺平面,这称为 17 个壁纸群。[76] 虽然在艺术和设计中很常见完全重复的铺砌形状,但在生物中却不容易找到。社群性黄蜂的纸蜂巢中的巢室,和蜜蜂的蜡蜂巢中的巢室,是众所周知的镶嵌例子。在动物中像硬骨鱼、爬行动物或穿山甲,或在植物中像蛇皮果这样的水果,都受到重叠鳞片或骨皮层的保护,这些鳞片或皮层大体上都是形状重复的单元,尽管它们实际上的大小通常是渐变的。在花㚏中,花格贝母的花瓣上有棋盘状的格纹。而矿物的结构则很好地示范了规律地重复的三维阵列。尽管已知的矿物有数十万种,但晶体中可能的原子排列方式却很少,因为受到晶体结构、晶系和点群所规限;例如对于三维空间中的 7 个晶格系统,在几何学上只可能有 14 种布拉维晶格。[77]
.jpg)






裂纹是在物料中形成的长形开口,以释放其所受到应力。当弹性物料均匀地拉伸或收缩时,它最终会达到其断裂强度,然后在各个方向突然断裂,产生 120°接头的裂缝,因此三个裂缝在一个节点处相遇。相反,当非弹性材料断裂时,则会形成直裂纹以释放应力。断裂之后,在同一方向上的应力将单纯地打开现有的裂缝;而直角方向上的应力则会产生新的裂缝,与旧裂缝成 90° 角。因此,裂缝的模式表明了该种物料是否具有弹性。[78] 例如在橡树树皮这样坚韧的纤维物料中,裂纹会如常形成以缓解应力,但因为受到树皮内的强弹性纤维束所阻,裂纹不会长得很长。由于每种树在细胞和分子层面上都有自己不同的结构,因此每种树的树皮都有自己独特的裂纹模式。[79]






动物的身体上经常呈现着各种图案模式:例如豹子和瓢虫身上有班点,神仙鱼和斑马身上有条纹。[80] 科学家尝试从进化学角度探索这些模式背后的成因:这些图案应该具有某些功能,令该种动物的后代增加了存活和繁殖的机会。动物图案的功能之一是伪装,[26] 例如,更难被察觉到的豹子会捕获到更多的猎物。另一个功能是发出信号,[27] 例如,瓢虫会较少被用肉眼捕食的掠食性鸟类攻击,如果它有大胆的警告颜色,并且还带有令人厌恶的苦味或毒性,或者模仿其他令人厌恶的昆虫。一只幼鸟可能会看到像瓢虫这样的昆虫并试图吃掉它,但它只会尝试一次,因为它很快便会吐出苦涩的虫子;而附近的其他瓢虫将不会受到捕食的干扰。年轻的豹子和瓢虫,继承了以某种方式产生斑点的基因,而得以生存。可是,虽然这些论点解释了为什么这些动物需要它们的图案,却仍没有解释这些图案本身是如何形成的。[80]

.jpg)




模式的形成机制
艾伦·图灵(Alan Turing)[17] 和后来的数理生物学家詹姆斯·莫瑞(James Murray)[81] 描述了一种自发产生斑点或条纹图案的化学机制:“反应-扩散系统”(reaction-diffusion system)。[82] 年轻有机体的细胞具有某些可以通过化学信号(形态发生素)开启的基因,从而导致某种组织结构的生长,例如深色的皮肤斑块。如果形态发生素无处不在,结果便是均匀的色素沉着,就像黑豹一样。但如果发生素分布得不均匀,就会产生斑点或条纹。图灵认为,形态发生素本身的产生可能存在着反馈机制。这可以导致形态发生素的数量在身体周围扩散时持续波动。此外,亦需要有另一个机制去产生驻波模式(而导致斑点或条纹):一种抑制性化学物质,它可以关闭形态发生素的产生,并且它本身能够比形态发生素更快地在体内扩散,从而形成一种“激活-抑制”机制组合。B-Z反应是这种机制的一个非生物学例子,即是一种所谓“化学振荡体系”(chemical oscillator)。[82]
后来的研究成功地创建了令人信服的图案模型,广泛地包括了例如斑马条纹、长颈鹿斑点、美洲虎斑点(被深色碎环包围的中等深色斑块)和瓢虫壳图案(不同的斑点和条纹的几何布局,见上一节的插图)等模式。[83] 理查德·普鲁姆的“激活-抑制模型”,从图灵的工作发展而来,使用六个变量来解释他们所观察到的九种基本羽毛内色素沉着的模式:从最简单的中央色素斑块,到同心斑块、条形、人字形、眼点、一对中心点、成对的点行,和点阵列。[84][85] 另一个更精细的模型模拟了普通珠鸡中复杂的羽毛图案,其中单个羽毛的特征是从羽毛基端的条形图案过渡到远端的一系列点图案。这种图案需要经由两个抑制信号所产生的化学振荡,在空间和时间上都有相互作用才能形成。[85]
在所谓“虎纹丛林”[86] 和“冷杉波浪”[87] 的植被景观中,图案模式可以基于各种物理上的原因而形成。虎纹丛林会出现在植物生长受降雨限制的干旱斜坡上:因为每个大致水平的植被条段都有效地收集了其上方裸露区域的雨水。[86] 而在山坡森林中,冷杉波浪会在风扰后,在再生长过程中发生:当一些树木被吹倒后,原先被它们遮蔽的树木就会暴露出来,进而更有可能受到破坏,因此树林的间隙往往会顺风扩大;而同时,在迎风面,年轻的树木在剩余的高大树木的风影的保护下得以生长。[87] 自然图案亦有时是由于动物的活动所造成,例如在美国西北部的美马冢和其他一些地区,有些图案似乎是由囊鼠多年的挖洞活动而形成的;[88] 而在纳米比亚出现的所谓“仙女环”,则似乎是由于沙漠白蚁群的相互竞争,以及沙漠植物之间对水的争夺,这两者的的相互作用而产生的。[89]
在具有活跃上层的永久冻土层中,会因每年的冻结和融化而产生所谓“图案地面”,形成圆形、网状、冰楔多边形、台阶和条纹。热收缩(负热膨胀)导致收缩裂缝的形成;到解冻时,水充满裂缝,在下一次冻结成冰时膨胀,并将裂缝扩大成楔形。这些裂缝可能会连接起来,而形成多边形和其他形状。[90]
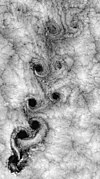
脊椎动物大脑上出现的裂隙模式是由“受限扩张”的物理过程引起的,这取决于两个几何参数:大脑皮层的相对切向扩张,和皮层的相对厚度。类似的脑回(峰)和脑沟(谷)模式已经在大脑模型中得到证实:模型从光滑的分层凝胶开始,在添加溶剂后,由外层(代表大脑的皮层)扩张所产生的压缩机械力可产生类似的裂隙模式。而在自然和实验上的观察中,这种表面折叠会随着大脑体积的增大而增加;这个观察亦得到了计算机模拟中的数值模型所支持。[91][92]









参见
参考资料
外部链接
Wikiwand in your browser!
Seamless Wikipedia browsing. On steroids.
Every time you click a link to Wikipedia, Wiktionary or Wikiquote in your browser's search results, it will show the modern Wikiwand interface.
Wikiwand extension is a five stars, simple, with minimum permission required to keep your browsing private, safe and transparent.