トップQs
タイムライン
チャット
視点
メガケイラ類
化石節足動物の分類群 ウィキペディアから

Remove ads
メガケイラ類[4][5](メガケイラるい、英: Megacheiran、学名: Megacheira[3])または巨鋏類[6](きょきょうるい)は、「大付属肢節足動物[7]」(great appendage arthropod[8]、大付属肢型節足動物[9])として知られる化石節足動物の分類群(綱)である[10]。同規的な体節に分かれた体と、手の造形に似た1対の「大付属肢[7][9]」(great appendage[11])をもつ[3]。ヨホイア、レアンコイリア、ハイコウカリス、フォルティフォルケプスなどが分類される[3][12][13]。

二十数種が記載され、確実の記録に限れば約5億年前のカンブリア紀から約4億5,000万年前のオルドビス紀の種類のみ含まれるが[2]、約4億3,000万年前のシルル紀と約4億年前のデボン紀に生息した、本群由来の可能性がある種類もわずかに知られている[注釈 1][1]。その分類学上の位置付けは議論的であり、節足動物の基部系統や鋏角類との関係性が特に注目される[10][14]。
Remove ads
名称
学名「Megacheira」はギリシャ語の「mega」(大きな)と「cheir」(手)の合成語で、本群の大きな手のように発達した大付属肢に因んで名付けられた[3]。総称は学名由来の「megacheiran」の他に「great appendage arthropod[8]」(大付属肢節足動物[7]、大付属肢型節足動物[9])がよく知られている[15][16][17]。ただしこの呼称は広い意味では「広義の大付属肢」をもつ本群以外の化石節足動物(ラディオドンタ類・イソキシス類など)もいくつか含まれる[18][19][20]ため、そこから区別できるように、本群は「short-great appendage arthropod」とも呼ばれている[1][17][21]。中国語では「大附肢類」(大附肢綱)と呼ぶ[22][23]。
形態
要約
視点


ほとんどのメガケイラ類は体長5cmに満たさない小型節足動物であり、例外的に大型のものとしてレアンコイリア(約7cm[24])、ヤウニク(9cm以上[25])とパラペイトイア(体の大部分は不明だが、前半身だけでも10cm以上ある[26])の種類のみ挙げられる。体は往々にしてエビのように細長く、付属肢(関節肢)は全て体の腹面にあり、数多くの体節は順に背甲に覆われる頭部と、十数節以上の背板に覆われる胴部という2つの合体節として区別される[27][13]。
頭部

 フォルティフォルケプスの頭部の背甲(cs)、眼(ey)、大付属肢(ba, pd)と脚(ca2-3)
フォルティフォルケプスの頭部の背甲(cs)、眼(ey)、大付属肢(ba, pd)と脚(ca2-3)



頭部(cephalon, head)は1枚の背甲(carapace)に覆われ、先頭は往々にして対になった眼をもつ。眼は複眼であり、1対もしくはレアンコイリア科において特徴的な2対で[28][29][24]、1対の場合は眼柄に突出したものがある[30][13]。一部の種類は、先頭背面に1枚の目立たない甲皮(anterior sclerite)と、その腹面から突出した1対の触角様の構造体(frontal protuberances)がある[3][31][30][10][13][32]。
頭部最初1対の付属肢は特徴的な大付属肢で(後述)、眼の直後、頭部の腹面から正面に張り出している。口は大付属肢の直後にあり、知られるものでは後ろ向きに開口して1枚の板状の構造体に覆われており、これは文献によりハイポストーマ(hypostome)、上唇(labrum)、もしくは両者の複合体(hypostome-labrum complex)と解釈される[24][10][33][32]。それ以降の後半部分は、後方ほど発達した数対の脚がある(後述)[27][13]。
付属肢の数と中枢神経系の構造(後述)[15][34][35]を基に、この頭部は一般に先節(眼とハイポストーマ/上唇)と第1-4体節(大付属肢と3対の脚)を含む合体節とされる[27]が、第3体節(第2対の脚)や第5体節(第4対の脚)まで含むとされる種類もある[13][32][2]。
大付属肢
→詳細は「大付属肢」を参照

メガケイラ類の最大の特徴である1対の大付属肢(great appendage[11])は、可動な関節をもつ、手のような造形をした捕獲用の関節肢である。5-6節の肢節をもつとされ、そのうち基部2節は上腕のような柄部(peduncle)で、残り3-4節は手のひらのようにまとまり、両部分の間に当たる関節(elbow-joint)は肘のように上向きに折り曲がる[36]。指のような爪(内突起 endite)は先端3-4節の各肢節の内側に1本ずつあり、隣接したものと噛合わせて”多重の鋏”(multi-chela[24])様の構造をなしているのが基本である[36][24][27]。一部の種類は爪の縁に鋸歯が並んでおり[25][20]、レアンコイリア目の場合、爪の先端には長い鞭毛(flagellum)がある[37][28][29][38][24][1][25][27]。
知られる限り、大付属肢の付け根は上唇/ハイポストーマ(口の開口)の直前にあり[37][24][33]、神経は中大脳のみに対応している[15][34]。そのため、この付属肢は第1体節由来で、少なくとも鋏角類の鋏角と他の節足動物の第1触角に相同であると考えられる(後述も参照)[15][10][27][34][13][33][20]。
メガケイラ類以外の化石節足動物における捕獲用の頭部付属肢、特にラディオドンタ類の前部付属肢とイソキシスの前端の付属肢まで含んだ「広義の大付属肢」[18][19][20]から区別できるように、この付属肢は「short-great appendage」("短い大付属肢"、SGA[21]、狭義の大付属肢 great appendage sensu stricto[20])とも呼ばれている[1][21][19][17][20]。
胴部


胴部(trunk)は体長の大部分を占める合体節で、種により11から20節以上の体節(胴節)が含まれる[31][27][13][20]。各胴節は背面から同規的な背板(tergite)に覆われ、背板の両縁が出張って肋部(tergopleura, pleurotergite)となる[24][36][13]。一部の種類は、各胴節の中央に目立たない腹板(sternite)をもつことが分かる[26]。最終胴節の末端は、種類により棘状か板状の尾節(telson)[37][36][16][39]、もしくは3枚の構造体でできた尾扇(tail fan)がある[3][13][20]。
脚

大付属肢、上唇/ハイポストーマと口以降に並ぶ十数対以上の付属肢は、基本として基部の原節(pasipod, protopod)・鰭(ひれ)状の外肢(exopod)・歩脚型の内肢(endopod)からなる二叉型の脚である[27]。一部の種類は、原節の内側に顎基(gnathobase, 内側の鋸歯状の内突起)[37][28][24][25]、外側基部に外葉(exite, 外側の目立たない葉状構造)[40]をもつことが分かる。これらの脚は原則として頭部に3対、各胴節に1対ずつある[27]が、頭部が2対か4対をもつとされる種類や[13][32]、最終数節の胴節が脚をもたない種類もある[36]。また、これらの脚は通常ではほぼ同形であるが、頭部の1対以上が短く特化した種類もある[26][24]。
神経系

C1/Ga: 大付属肢神経、C2-4: 第2-4頭部付属肢神経、Cg: 頭部神経節、Cn: 連合神経、Dc: 中大脳、Of: 食道孔、On: 視神経、Op1-2: 第1と第2視神経網、Pc: 前大脳、T1-7: 第1-7胴節/胴部付属肢神経、Tc: 後大脳、Tg: 胴部神経節
神経系はアラルコメネウス(澄江動物群[15]、Pioche Formation[34] と Marjum Formation[34])とレアンコイリア(Kaili Formation[35])のみから知られ、視神経と中枢神経系が発見される[15][34][35]。1対の視神経は前大脳の両前方に連結し、それぞれ途中から二股に分かれて2対の眼に対応する[15][34]。視神経網(optic neuropil)のうち、第1と第2視神経網はそれぞれ前大脳の外側と内側に配置される[15][35]。頭部は先節と第1-4体節由来とされる5節の神経節(ganglion)を含め、前3節は順に一般的な節足動物と同様な脳神経節(先節の前大脳 protocerebrum・第1体節の中大脳 deutocerebrum・第2体節の後大脳 tritocerebrum)とされ、前大脳は視神経、中大脳は大付属肢、後大脳以降の3節は頭部3対の脚に対応する[15][34][35]。前大脳の先頭の一部は更に「prosocerebrum」として区別でき、これは内側1対の眼と上唇の神経に対応する[35]。消化管に貫通される食道孔(oesophageal foramen)は、前大脳の直後から4節目の神経節まで大きく開いている[15][34]。胴部の腹神経索(ventral nerve cord)は第1-8胴節にかけて各胴節と付属肢に対応する神経節が並んでいるが、第9胴節以降では第8胴節から伸ばした連合神経(connectives)のみ配置され、神経節はない[15][34]。
このような神経系、特に2対に分かれた視神経・脳の外側に配置される1対の視神経網・集約した頭部神経節・前大脳の直後まで広げた食道孔などの性質は、節足動物の中で鋏角類に最も似ている[15][41]。なお、前大脳の先頭(prosocerebrum)が明確に区別されるのは現生節足動物の系統群に似ておらず、むしろラディオドンタ類[42]などの基盤的な節足動物の脳神経節を思わせる[35]。
消化系
消化系(消化管)は頭部の腹側で後ろ向きに開口する口から始め、上向きに曲がり返した咽頭を介して腸に連結する[37][28][1]。腸は頭部の咽頭から胴部の後端まで走り、レアンコイリア、Oestokerkus とヤウニクの場合、胴節に対応する消化腺(digestive gland, diverticula, 中腸腺 midgut gland)が中腸で対になって配置されることが分かる[43][38][44][45][25]。肛門は尾節と最終胴節の腹面の境目に開口する[37][25]。
Remove ads
生態

ヤウニク

フォルティフォルケプス(左奥)と Sklerolibyon(右手前)
海底を泳ぐメガケイラ類の生態復元図

メガケイラ類は全般的に遊泳性(nektonic)から遊泳底生性(nektobenthic, 遊泳性に近い底生性)で、捕食者もしくは腐肉食者であったとされる[46]。脚の外肢で海中を泳ぎ、捕獲用の大付属肢で餌を捕らえたと考えられる[46]。例えばヨホイアの大付属肢は、甲殻類のシャコの捕脚のように、瞬発に突き出して獲物を捕獲できたと推測される[36]。レアンコイリア目の種類に見られる鞭毛をもつ大付属肢は、捕食と感覚の役割を兼ね備えたと考えられる[37][25]。脚に顎基をもつ種類は、それを用いて餌を粉砕できたと考えられる[37][24]。一部の種類は発達した消化腺をもつことにより、餌を効率よく蓄積・消化できたことも示唆される[43][29][45]。
レアンコイリアの場合、幼生は甲殻類のノープリウス幼生を彷彿とさせ、胴節が成体より少なく、大付属肢の形態もやや異なることが分かる[21][17]。これにより、少なくとも本属のメガケイラ類は幼生と成体のニッチ(生態的地位)が分化しており[21]、体節を増やしながら成長した(増節変態を行った)と考えられる[17]。
分布と生息時代
要約
視点


ほとんどのメガケイラ類の化石標本は北アメリカと中国におけるカンブリア紀の堆積累層で見つかり、中でもカナダのバージェス頁岩(バージェス動物群)と中国雲南省の Maotianshan Shale(澄江動物群)から特に多く知られている[47][3][24][46]。ほとんどの属は1つの堆積累層のみに分布するが、レアンコイリアとアラルコメネウスは例外的に広い分布域をもつ[34]。
もしイギリスの Herefordshire Lagerstätte による Enalikter aphson [1]とドイツのフンスリュック粘板岩による Bundenbachiellus giganteus [1]はメガケイラ類であれば、本群の生息時代はカンブリア紀を超えて、シルル紀とデボン紀前期にも及ぶとなる[1]。
メガケイラ類が発見される堆積累層は次の通り。本群としての本質が不確実性をもつ記録は「*」で示す。同定または独立種としての有効性が不確実の記録は「?」で示す。
- カンブリア紀第三期(約5億2100万 - 5億1400万年前)
- Qingjiang biota(中国、湖北省、約5億1,800万年前):Alalcomenaeus sp.[51]、Leanchoilia sp.[51]、レアンコイリア科新種A[51]
- カンブリア紀第四期(約5億1400万 - 5億900万年前)
- カンブリア紀ウリューアン期(約5億900万 - 5億450万年前)
- Kaili Formation(Kaili Biota、中国、貴州省、約5億800万年前[53]):Leanchoilia robisoni[44]、Leanchoilia sp.[35]
- Spence Shale(アメリカ、ユタ州):Leanchoilia? hanceyi[54][55]、Leanchoilia superlata?[55]、Yohoia utahana[56]
- バージェス頁岩(バージェス動物群、カナダ、ブリティッシュコロンビア州、約5億1,000万 - 5億500万年前[57]):Actaeus armatus[58]、Alalcomenaeus cambricus[58][37]、Leanchoilia persephone[58][29]、Leanchoilia protogonia[58][29]、Leanchoilia superlata[47][24]、*Worthenella cambria[59]、Yohoia tenuis[47][36]、Yawunik kootenayi[25]
- Stephen Formation(カナダ、ブリティッシュコロンビア州):*Kootenichela deppi[31]
- カンブリア紀ドラミアン期(約5億450万 - 5億50万年前)
- Wheeler Shale(アメリカ、ユタ州):Kanoshoia rectifrons[60]、Leanchoilia cf. L. illecebrosa[60]
- Marjum Formation(アメリカ、ユタ州、約5億200万年前):Alalcomenaeus sp.[34]、Leanchoilia? sp. cf. protogonia[55]
- カンブリア紀フロンギアン期(約4億9,700万 - 4億8,540万年前)
- オルドビス紀カティアン期(約4億5,300万 - 4億4,20万年前)
- Beecher's Trilobite Bed(アメリカ、ニューヨーク州):Lomankus edgecombei[2]
- シルル紀(約4億4,370万 - 4億1,600万年前)
- Herefordshire Lagerstätte(イギリス、約4億3,000万年前[62]):*Enalikter aphson[1]
- デボン紀プラギアン期 - エムシアン期(約4億1,080万 - 3億9,330万年前)
- フンスリュック粘板岩(ドイツ、約4億800万 - 4億年前):*Bundenbachiellus giganteus[63][1]
Remove ads
大付属肢の対応関係
要約
視点
メガケイラ類の大付属肢の由来や他の節足動物の付属肢との対応関係(相同性)は、その類縁関係(後述)を推測するのに重要視される基準の1つであるため、課題として注目を集め、多くの議論をなされていた[10]。これは文献により前大脳性(先節由来)[64][65]・中大脳性(第1体節由来)[12][66][67][15][13]・後大脳性(第2体節由来)[68][69]・ラディオドンタ類(アノマロカリスなど)の前部付属肢に相同[64][12][66][70][67][71][72][13][20][65][73]・鋏角類(クモ・サソリ・カブトガニなど)の鋏角に相同[12][66][71][72][15][27][33][13]・大顎類(多足類・甲殻類・六脚類)とArtiopoda類(三葉虫・光楯類など)の第1触角に相同[12][66][70][67][15][74]・甲殻類の第2触角に相同[3]まで、数々の解釈を与えられた[10]。
2000年代から2010年代初期にかけて、大付属肢の対応関係がこうして諸説に分かれるのは、それを直接的に証明できる神経解剖学的証拠(大付属肢神経と脳神経節の構造)が長らく不明であったことが大きな理由である[10]。しかし2010年代後期以降では、メガケイラ類の脳神経節(#神経系を参照)が判明した[15][34][35]ことにより、大付属肢は中大脳性/第1体節由来で、すなわち鋏角と第1触角に相同する説が最も広く認められるようになった[10]。
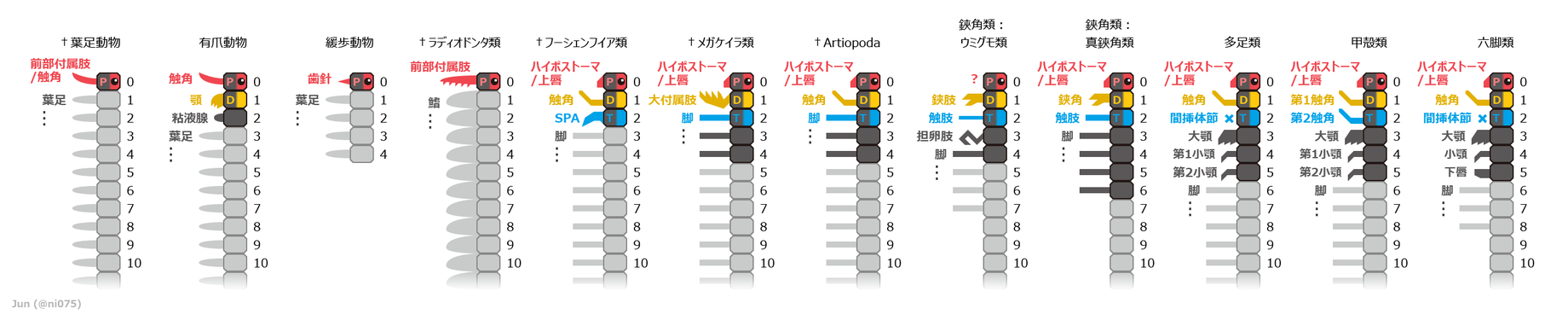 2010年代後期で広く認められる、様々な汎節足動物における体節と付属肢の対応関係[10]。
2010年代後期で広く認められる、様々な汎節足動物における体節と付属肢の対応関係[10]。

大付属肢に対して提唱された様々な対応関係の詳細(根拠・経緯・有効性など)は、次の通りに示される[10]。
- 中大脳性/第1体節由来[12][66][67][15][13][35]:鋏角類の鋏角、Artiopoda類と大顎類の第1触角に相同[12][66][70][67][71][72][15][74][27][33][13][35]
- メガケイラ類(アラルコメネウスとレアンコイリア)で見つかった脳神経節の構造から直接に証明される[15][34][35]、最も有力で広く認められる対応関係[10]。
- 上述の神経解剖学的証拠以外にも、中大脳性付属肢のように眼と上唇/ハイポストーマの間にあること[10][33]、および鋏角との類似性(同じく捕獲用の前端の付属肢で、柄部と鋏に分化した前後の肢節をもつなど[12][75][36])が根拠として挙げられる。
- 後大脳性/第2体節由来[76][68][69]:甲殻類の第2触角に相同[3]

- 一部のメガケイラ類(フォルティフォルケプス、Kootenichela など)が大付属肢の直前に触角様の構造体をもち、それを中大脳性の第1触角と解釈することによって提唱された対応関係[3][76][68][69]。これにより、触角の直後にある大付属肢は必然的に後大脳性/第2体節由来となり、触角をもたないメガケイラ類は単にそれを二次的に退化したと考えられる[69]。
- 鋏角に相同する説に踏まえてこの対応関係を提唱する文献もある[76]が、これは中大脳性/第1体節由来の鋏角を、後大脳性/第2体節由来と誤解釈した古典的な見解(鋏角類#体節と付属肢の対応関係を参照)に基づいたため、無効とされる[10]。
- 上唇/ハイポストーマとの位置関係や神経解剖学的証拠に対応しないことと、触角様の構造体が形態的に中大脳性付属肢である触角として認められにくいことにより否定的とされる[10]。
- ラディオドンタ類の前部付属肢に相同[64][12][66][70][67][71][72][13][20]

- 前部付属肢との類似性(捕獲用の前端の付属肢・柄部に分化した基部の肢節・内突起/爪に鋸歯/分岐があるなど[12][72][24][20])、およびラディオドンタ類と真節足動物の中間型生物の形態(例えばキリンシアとフェンゼンギアは、大付属肢と同じ位置にある前端の付属肢が前部付属肢的である)を基に提唱される対応関係[20]。
- この対応関係は、主に両付属肢を中大脳性/第1体節由来とする説に踏まえて提唱される[66][70][67][13]。少数派であるが、両付属肢を前大脳性/先節由来とする説[64][65]、もしくは前部付属肢のみ前大脳性/先節由来とする説に踏まえて提唱する文献もある[20]。
- 2010年代後期では、メガケイラ類(アラルコメネウス)とラディオドンタ類(ライララパクス)で見つかった脳神経節の構造の違い(大付属肢は中大脳性/第1体節由来[15][34]、前部付属肢は前大脳性/先節由来とされる[42])により、この相同性は多くの文献に否定的とされるようになった[42][77][10][78]。ただし2020年代以降では、ラディオドンタ類の神経解剖学的証拠の見直し(前部付属肢を中大脳性/第1体節由来とする)により再び相同とされ[13][20][73]、また上述の通りに対応する体節と脳神経節が別だとしても相同の可能性がある(ホメオティック遺伝子変異を通じて相同の付属肢を別の脳神経節/体節に反映させた)という、両付属肢の相同性を支持する文献もある[20]。→「ラディオドンタ類 § 前部付属肢の対応関係」、および「キリンシア § 前端の付属肢の相同性と進化」も参照
- 前大脳性/先節由来:他の節足動物の上唇/ハイポストーマに相同[64]
- 前述の前部付属肢に相同する説と、前部付属肢の前大脳性説を同時に踏まえて提唱される対応関係[64][65]。
- メガケイラ類が上唇/ハイポストーマと大付属肢を同時にもつことと、神経解剖学的証拠に対応しないことにより否定的とされる[10]。
他にもイソキシス類の前端の付属肢、フォルフェクシカリスとオッカカリスの前端の付属肢、フーシェンフイア類のSPAs(specialized post-antennal appendages)など、いわゆる「広義の大付属肢」に含まれる、他の化石真節足動物における捕獲用の付属肢との対応関係も議論をなされていた[64][66][70][67][68][69][10][65]。SPAsの大付属肢との相同性は否定的で、一般に後大脳性/第2体節由来[79][80]もしくは第3体節由来[81]とされる。それ以外のものに関しては、脳神経節と上唇/ハイポストーマの構造が未だに不明のため、対応関係は不確かである[10]。
→詳細は「大付属肢 § メガケイラ類以外の "大付属肢"」を参照
Remove ads
分類
要約
視点
系統位置
| ||||||||||||||||||||||||||||||
| メガケイラ類の系統位置に対する主流の2つの解釈[14]。 A:基盤的な真節足動物説、B:基盤的な鋏角類説 |
最初に記載されたメガケイラ類、例えばレアンコイリアとヨホイアは20世紀前期では鰓脚類の甲殻類と解釈された[47]。20世紀後期では、これらの化石節足動物は(鰓脚類とは別系統の)1つの分類群を表しているものと判明し、Bergström 1992 で暫定的に「大付属肢節足動物」(great appendage arthropod)と総称され[8]、Hou & Bergström 1997 により正式にMegacheira(綱)と命名されたが、当時は依然として甲殻類の近縁と考えられた[3]。21世紀以降から研究が進み、一般に甲殻類とは別系統の節足動物とされるようになった[19][10]。
メガケイラ類の系統位置は議論的であり、他の節足動物に対する単系統性もしばしば疑問視されている(#下位分類も参照)[64][12][72][36][82][69][25][83][13][84]。その系統位置に対する解釈は、主に「基盤的な鋏角類」[12][76][75][72][36][15][74][25][33][20][85][73][2]と「基盤的な真節足動物」[64][82][31][68][69][1][86][83][87][13][84][88]という2説に分かれ[89][77][10][14][41]、ラディオドンタ類との関係性も議論の対象となった[64][12][72][89][36][10][13][20]。
2000年代~2010年代前期の展開






| ||||||||||||||||||||||||||||||||||||
| ラディオドンタ類とメガケイラ類(太字、青枠)を基盤的な鋏角類とし、後者の中でハイコウカリスをより鋏角類に近いとする系統仮説[12][72][36]。なお、このような位置関係は系統解析に支持されず、2010年代以降では否定的である[10]。 |
Budd 2002 では、大付属肢の前大脳性/先節由来説(ほとんどの文献に否定される対応関係[10])を基に、メガケイラ類はラディオドンタ類に類縁の基盤的な真節足動物と考えられた[64]。一方、Chen et al. 2004 と Cotton & Braddy 2004 をはじめとして、大付属肢と鋏角の類似点(捕獲用の前端の付属肢・柄部と鋏に分化した前後の肢節をもつなど)が注目され、メガケイラ類を基盤的な鋏角類とする系統仮説が提唱された[12][76][75]。特にメガケイラ類の中で、ハイコウカリスは爪3本の大付属肢が鋏角に最も似ているとされ、他のメガケイラ類より鋏角類に近く、メガケイラ類全体は鋏角類に至る側系統群と考えられた[12][75][72][36]。
Chen et al. 2004 と Haug et al. 2012 では鋏角類の他に、ラディオドンタ類の前部付属肢と大付属肢の類似点も示されており、ラディオドンタ類はメガケイラ類に、すなわち鋏角類にも近縁と考えられた[12][72][36]。これらの説を統合し、節足動物の中でラディオドンタ類とメガケイラ類は順番ずつ鋏角類の初期系統のみから分岐するとされ、肢節数と内突起/爪の数を減らしながら、前部付属肢(十数節以下、内突起5本以上)は大付属肢(6節以下、爪4-3本)に、大付属肢は鋏角(3節以下、爪2本)に進化すると解釈された[12][71][72][36]。更に Haug et al. 2012 では、フルディア科(フルディア、ペイトイアなど)のラディオドンタ類は前部付属肢が比較的少数(10節以下)の肢節と数本の爪らしき内突起をもつことにより、他のラディオドンタ類よりメガケイラ類/鋏角類の系統に近く、ラディオドンタ類全体はメガケイラ類+鋏角類の分岐に至る側系統群という説も提唱された[36]。
また、メガケイラ類のパラペイトイアは Budd 2002 と Chen et al. 2004 の両方においても「メガケイラ類の大付属肢をもつラディオドンタ類」と解釈され、ラディオドンタ類とメガケイラ類の中間型生物とされた[64][12]。

灰色:確実な形質、赤色:不確実なラディオドンタ類的形質、破線:不明部分
2010年代以降の展開
→「ラディオドンタ類 § 系統関係」、および「鋏角類 § 化石節足動物との関係性」も参照
しかしラディオドンタ類に関しては、2010年代以降からこれらの真節足動物との類縁関係が徐々に否定され、外骨格が進んでいない胴部や単調な脳神経節ど多くの祖先形質により、真節足動物自体より早期に分岐する基盤的な節足動物として広く認められるようになった(ラディオドンタ類#分類を参照)[42][77][90][10]。加えて、Cong et al. 2014 によって示される前部付属肢の前大脳性/先節由来(すなわち中大脳性/第1体節由来の大付属肢に非相同)の可能性も、ラディオドンタ類とメガケイラ類の類縁関係に疑問を掛けている[42][77][10]。また、パラペイトイアも中間型生物としての情報性を失い、ラディオドンタ類的形質が否定的に評価され、単にメガケイラ類の見間違いとして広く認められるようになった(パラペイトイア#系統関係を参照)[91][82][31][69][31][68][90][46][13]。
こうしてメガケイラ類とラディオドンタ類の類縁関係は2010年代以降から否定的になり[10]、ハイコウカリスを他のメガケイラ類より鋏角類に近いとする説も、ほとんどの系統解析に支持されなかった[82][31][74][68][69][1][25][86][83][87][13][84][20][85][73]。しかしメガケイラ類自体と鋏角類の類縁関係は議論の余地が残され、前述の大付属肢と鋏角の類似性だけでなく、後に判明した鋏角類的な脳神経節(#神経系を参照)[15][41]と退化的な上唇/ハイポストーマ[33]、およびいくつかの系統解析結果[74][15][25][92][33][20][85][73]にも示唆される。他方、この類縁関係を認めず、メガケイラ類を鋏角類・Artiopoda類(三葉虫、光楯類などの系統群)・大顎類(多足類、甲殻類、六脚類の系統群)より早期に分岐した基盤的な真節足動物とする解析結果も少なからぬ挙げられる[82][31][68][69][1][86][83][87][13][84][88]。もしメガケイラ類は基盤的な真節足動物であれば、前述の鋏角類的性質は収斂進化した同形形質とされ[10][41]、ラディオドンタ類などの基盤的な節足動物を思わせる前大脳の prosocerebrum の構造(#神経系を参照)も、文献によりこの系統位置を裏付ける証拠とされる[35]。
下位分類
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 基盤的な鋏角類とされるメガケイラ類を単系統群とする系統解析の例(Zeng et al. 2020)[20] |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 基盤的な真節足動物とされるメガケイラ類を単系統群とする系統解析の例(Zhai et al. 2019)[87] |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| 基盤的な真節足動物とされるメガケイラ類(青枠)を側系統群とする系統解析の例(Aria et al. 2020)[13] |
| |||||||||||||||||||||||||||||||||||||||
| 基盤的な真節足動物とされるメガケイラ類(*)を多系統群とする系統解析の例(Aria & Caron 2017)[83] |
メガケイラ類の単系統性は不確実であり[10]、2010年代以降の系統解析だけでも、文献によっては単系統群[74][86][92][87][20][85][2]、側系統群[36][82][68][69][1][92][13]、更に多系統群[83][84]とされる場合もある。これは系統解析がなされる頃において、一部の種類、特に本群の中で比較的に特異な形態を有するフォルティフォルケプスとジェンフェンギアの多くの形質が不明確であったことが大きな理由とされる[83]。少なくともレアンコイリア科(レアンコイリア、アラルコメネウス、ヤウニクを含んだ分類群)の単系統性は広く認められ、ヨホイア、ハイコウカリスと単系統群(Cheiromorpha[25])になる説も比較的に多くの支持を得られている[25][86][83][92][93][84]。




蠕虫様の細長い胴部をもつ Enalikter aphson に関しては、メガケイラ類どころか、節足動物ですらなく、むしろ多毛類(ゴカイ類)の環形動物ではないかという異説があった[94]。しかしこの知見は、後に本種の節足動物的性質(関節肢や曲がり返した消化管など)を補足した反発文献によって否定された[95][96]。しかし本種はよく似た Bundenbachiellus giganteus と並んで、メガケイラ類的性質は依然として一部の文献に疑問視されている[25]。Kootenichela deppi として記載された化石標本は、複数種の節足動物に由来で、特にその模式標本はメガケイラ類ですらない別系統のものと疑わしく見受けられる[97][13]。甲殻類として記載され[61]、更にいくつかの文献に基盤的な甲殻類[98][99][100]と考えられた Oelandocaris oelandica は、実はレアンコイリア科のメガケイラ類ではないかという説もある[25][19][86]。
他にも、Tanglangia、フォルティフォルケプス、ジェンフェンギアという3属は同種による別の成長段階[46]、レアンコイリア属のいくつかの種は同種による性的二形[29]、Actaeus は単に保存状態が良くないアラルコメネウス[37]など、広く認められる意見ではないが、別種として記載されたメガケイラ類の化石標本の中で、実は同種によるものがあるではないかと疑問視される場合もある。
2020年まででは、不確実の記録をも含めて次の19属二十数種のメガケイラ類が正式に命名される(属より上位の分類群は太字、不確実の記録「?」で示される)。
- メガケイラ綱 Megacheira [3]
- (属)Tanglangia [50]
- ジェンフェンギア科 Jianfengiidae [13]
- フォルティフォルケプス属 Fortiforceps [3]
- ジェンフェンギア属 Jiangfengia [49]
- パラペイトイア(パラペユトイア)属 Parapeytoia [26]
- (属)Sklerolibyon [13]
- Sklerolibyon maomima [13]
- ?(科)Kootenichelidae [31]
- ?(属)Kootenichela [31]
- ? Kootenichela deppi [31]
- ?(属)Worthenella [59]
- ?(属)Kootenichela [31]
- (階級なし)Cheiromorpha [25]
- ハイコウカリス属 Haikoucaris [12]
- Haikoucaris ercaiensis [12]
- レアンコイリア目 Leanchoiliida [101]
- ?(科)Enaliktidae [1]
- レアンコイリア科 Leanchoiliidae[11](=Actaeidae [106][28]、=アラルコメネウス科 Alalcomenaeidae [106][28])
- ヨホイア目 Yohoiida [108]
- ハイコウカリス属 Haikoucaris [12]
Remove ads
類似群
→「大付属肢 § メガケイラ類以外の "大付属肢"」も参照
 フォルフェクシカリス
フォルフェクシカリス オッカカリス
オッカカリス







メガケイラ類に似た特徴をもつカンブリア紀の化石節足動物はいくつか挙げられる。フォルフェクシカリス(Forfexicaris)[109]とオッカカリス(Occacaris)[109]という、かつてメガケイラ類と解釈され[109]、体が二枚貝様の甲皮に覆われた不明点の多い節足動物は、大付属肢によく似た1対の捕獲用の付属肢がある[20]。イソキシス(Isoxys)の一部の種類も、前端の付属肢に大付属肢を思わせる性質が見られる[18][97][10][20]。Kiisortoqia [91]とキリンシア(Kylinxia)[20]は前端の付属肢がラディオドンタ類の前部付属肢によく似ているが、胴部の構成はむしろメガケイラ類に似て、特に後者の前端の付属肢は大付属肢に近い性質もわずかにあわせもつ(キリンシア#他の節足動物との比較を参照)[20]。これらの節足動物は、ほとんどがメガケイラ類とは別系統扱いされており[74][25][83][13][20]、その類似性は、単なる祖先形質の名残もしくは収斂進化の結果と考えられる[74][10][20]。
Remove ads
脚注
関連項目
外部リンク
Wikiwand - on
Seamless Wikipedia browsing. On steroids.
Remove ads