Rondwormen
grote stam van zeer algemeen voorkomende wormen Van Wikipedia, de vrije encyclopedie

Rondwormen (Nematoda) zijn een grote groep (stam) van zeer algemeen voorkomende wormen. Er zijn meer dan 25.000 beschreven soorten van de naar schatting 10 miljoen bestaande soorten.[2] Ook de aaltjes behoren tot de rondwormen. Plantenparasitaire rondwormen worden meestal aaltjes genoemd. Nematologie is de wetenschap die de nematoden bestudeert. De term nematoda komt uit het Grieks en betekent “draad-achtig”. Ongeveer 90% van de rondwormen bevindt zich in de bovenste 15 cm van de bodem. Rondwormen kunnen ook een belangrijke rol spelen in de stikstofkringloop door stikstofmineralisatie.[3]
| Rondwormen Fossiel voorkomen: Proterozoïcum[1] — heden | ||||||
|---|---|---|---|---|---|---|
 | ||||||
| Een rondworm die soja-bonen aantast (met ei). | ||||||
| Taxonomische indeling | ||||||
| ||||||
| Stam | ||||||
| Nematoda Rudolphi, 1808 | ||||||
| Afbeeldingen op Wikimedia Commons | ||||||
| Rondwormen op Wikispecies | ||||||
| (en) World Register of Marine Species | ||||||
| ||||||






Rondwormen hebben zich met succes aangepast aan bijna elk ecosysteem: van zee (zout) tot zoet water, in bodems van de poolgebieden tot de tropen van hoog tot laag. Ze zijn alomtegenwoordig in zoetwater-, zee- en terrestrische omgevingen, waar ze vaak andere dieren in aantallen overtreffen in zowel individuën als soorten en worden gevonden op locaties zo divers als bergen, woestijnen en troggen. Ze zijn te vinden in elk deel van de lithosfeer, [4] zelfs op grote diepte, van 0,9 tot 3,6 km onder het aardoppervlak in goudmijnen in Zuid-Afrika.[5][6][7][8][9] Ze vertegenwoordigen 90% van alle dieren op de oceaanbodem.[10] De hoogste dichtheden rondwormen zijn waargenomen in de toendra en boreale bossen.[11] De vele parasitaire soorten zijn pathogenen van de meeste planten en dieren. Een derde van de geslachten komt voor als parasiet van gewervelden; ongeveer 35 rondwormsoorten komen bij mensen voor.[12]
Veel rondwormen eten bacteriën, schimmels en protozoa. Ze kunnen de bacteriepopulatie en de samenstelling effectief reguleren en kunnen tot 5000 bacteriën per minuut eten.[3] Andere soorten leven als parasiet en veroorzaken plantenziekten zoals aardappelmoeheid, tasten insecten, naaktslakken aan of zitten in het darmkanaal van zoogdieren zoals de spoelwormen of de aarswormpjes en worden in het laatste geval dan vaak als draadworm aangeduid. Entomopathogene rondwormen worden ingezet ter bestrijding van schadelijke insecten,[13] zoals de inzet van Steinernema feltiae bij de bestrijding van de eikenprocessierups.[14] en het gebruik van Phasmarhabditis hermaphrodita tegen naaktslakken.
Sommige rondwormen kunnen wel 8 meter lang worden (Placentonema gigantissima),[15][16] maar de meeste rondwormen zijn voor het blote oog onzichtbaar en alleen door een microscoop zichtbaar. Ze zijn dan 5 tot 100 µm dik en 0,1 tot 2,5 mm lang.[3] Per m² bodem kunnen wel een miljoen rondwormen leven.
Rondwormen kunnen aangetast worden door onder andere nematofage schimmels, virussen, sporozoën, amoeben, ciliaten, tardigraden, colembolen, oligochaeten en bacteriën.[17]
Genoom
Samenvatten
Perspectief
De soort Caenorhabditis elegans wordt gebruikt als modelorganisme. Het was de eerste meercellige eukaryoot waarvan het genoom volledig in kaart werd gebracht in 1998 en bestaat uit zes chromosomen met ongeveer 100 Mbp en 20.000 genen.[18] Ook is van C. elegans de ontwikkelingsbiologie van elke cel bepaald en elke neuron in kaart gebracht. Het ook uit zes chromosomen bestaande genoom van Caenorhabditis briggsae is in 2003 in kaart gebracht en is ook ongeveer 108 Mbp groot met 22.000 genen.[19][20][21] Caenorhabditis nigoni is nauw verwant aan Caenorhabditis briggsae en heeft een genoom dat 129 Mbp groot is met 29.167 coderende genen.[22][22]
Ook is het genoom van Meloidogyne incognita in 2008 volledig gesequencet en bestaat uit 86 Mbp.[23] Het andere modelorganisme ter vergelijking met Caenorhabditis elegans is Pristionchus pacificus, waarvan het genoom ook volledig in kaart is gebracht, die op zijn beurt weer is vergeleken met Parapristionchus giblindavisi.[24] Het genoom van Trichinella spiralis is eveneens volledig in kaart gebracht en in 2011 gepubliceerd.[25] Dit genoom is 58,55 Mbp basenparen groot met een schatting van 16.549 genen.[26] Het Trichinella spiralis-genoom is het enige rondwormgenoom met het epigenetische mechanisme van DNA-methylatie,[27] Het genoom van Necator americanus is in 2014 gesequencet en geanalyseerd.[28] Het genoom is 244 Mbp groot en bestaat uit 19.151 eiwitcoderende genen.
Fossielen
Samenvatten
Perspectief
Rondwormen hebben geen lichaamsdelen die gemakkelijk kunnen worden geconserveerd gedurende geologische tijdsperioden en die relatief gemakkelijk kunnen worden geïdentificeerd in sedimenten en sedimentair gesteente. Daarom is hun fossielenbestand erg fragmentarisch en klein in relatie tot hun brede verspreiding en diversiteit, en is het beperkt tot specifieke conserveringsvormen of sedimentaire facies.[29][30]
De best bewaarde rondwormfossielen werden gevonden in barnsteen uit het Krijt en het Tertiair, waarvan de oudste uit het Onder-Krijt in Libanon stammen.[29][31][32] De vroegste directe fossielenbestanden (lichaamfossielen) zijn afkomstig uit de beroemde Rhynie Chert in Schotland en zijn ook het oudste directe bewijs voor parasitaire rondwormen. Ze bestaan uit eieren, verschillende larvenstadia en volwassen individuen ontdekt in de subhuidmondjesholte ("respiratory cavity") van de vroege landplant Aglaophyton major en in 2008 onder de naam Palaeonema phyticum beschreven.[29] Sedimentstructuren met sporenfossielen van vrij levende mariene, infestale rondwormen werden gevonden in kleisteen uit het Ordovicium in de Hubei Provincie in China.[30]
In het Laagland van Kolyma in Siberië werden in juli 2018 door Russische wetenschappers tijdens bodemonderzoek in de Pleistocene Parken rondwormen gevonden die respectievelijk ongeveer 42.000 en 32.000 jaar in de permafrost bodem zijn bewaard. Ondanks het feit dat ze waren bevroren sinds het Pleistoceen zijn twee exemplaren, genaamd Panagrolaimus aff. detritophagus en Plectus aff. parvus succesvol tot leven gewekt. (aff. is een afkorting van het Latijnse affinis en wil zeggen dat het nauw verwant is aan).[33] Terwijl er in de wetenschappelijke wereld nog twijfels bestaan over de juistheid van deze bevindingen, wordt er algemeen aangenomen dat het theoretisch niet onmogelijk is dat deze dieren na een dergelijk lange periode terug tot leven gewekt kunnen worden.
Lichaamsbouw
Samenvatten
Perspectief
Rondwormen zijn ongesegmenteerde, ronde, oermondige dieren met een compleet spijsverteringsstelsel. Ze behoren daarmee tot de meest eenvoudige dieren die toch een compleet verteringsstelsel hebben. Ze hebben geen hart- en vaatstelsel of ademhalingsstelsel en zijn dus aangewezen op diffusie om zuurstof binnen te krijgen en te transporteren.[34] De dwarsdoorsnede van rondwormen is rond en bestaat uit een kop, middenstuk en staart. De kop is radiaal symmetrisch en de rest van het lichaam is tweezijdig symmetrisch.
De kop heeft in veel gevallen uit een geheel bestaande 'kopschermen' die rond de mond naar buiten uitstralen.[35] Bij de basis van de lippen zitten amfiden. Dit zijn de belangrijkste reukorganen van een rondworm. Elke amfide bestaat uit twaalf sensorische zenuwcellen en met trilhaartjes bezette dendrieten.
Er zijn rondwormen met een of twee ogen, waarbij er meestal twee ogen aanwezig zijn, zoals bij Chromadorina bioculata, Oncholaimus vesicarius, Enoplus anisospiculus en Deontostoma californicum. De vrouwelijke Mermis nigrescens-rondworm heeft slechts een oog, waarvan de bouw afwijkt van de ogen van rondwormen die er twee hebben. Deze ogen bevatten melanine als schaduwpigment, terwijl dat bij Mermis nigrescens oxyhemoglobine bevat. Het oog van Mermis nigrescens bestaat uit een holle cilinder.[36] en het rode pigment binnenin bleek hemoglobine te zijn.[37] Deze hemoglobine, meestal als oxyhemoglobine, is dicht geconcentreerd in kristallijne vorm.[36] De cilinder bleek een ocellus met een soort hoornvlies te zijn.[38]
Op het lichaam komen vaak groeven, ribbels (synlofe=gebied met ribbels), borstelharen of andere onderscheidende structuren, zoals striae voor.[39]
Bij de geslachten Trichuris en Capillaria komen bacillaire banden voor. Dit zijn gespecialiseerde rijen van in de lengterichting liggende cellen. Ze bestaan uit kliercellen en een ander type cellen en worden gevormd door de hypodermis. De kliercellen hebben poriën tot in de huid. De functie van de bacillaire banden is onbekend. Hun buitengewone structuur duidt erop dat de kliercellen mogelijk een rol spelen in osmotische of ionenregulatie. De andere cellen spelen mogelijk een rol bij de vorming van de huid en de opslag van voedsel.
De 'huid' van een rondworm bestaat uit een syncytium of een enkele laag cellen en is bedekt met een dikke collagene cuticula of cuticulum. De cuticula heeft vaak een complexe structuur en kan uit twee tot vier verschillende lagen bestaan. Van binnen naar buiten:
- De binnenste vezellaag bestaat uit diagonaal tegenovergestelde vezels die in tegengestelde richtingen lopen. Deze laag draagt het meest bij aan de sterkte en elasticiteit van de cuticula.
- De matrixlaag heeft een minder gedefinieerde structuur.
- De corticale laag bestaat uit collageen.
- De buitenste epicuticula is lipide-houdend en wordt bij sommige rondwormgeslachten ook nog bedekt door een lipidelaag.
,Caenorhabditis_elegans_(left)_body_wall_muscle.jpg)
Onder de 'huid' ligt een laag van in de lengte liggende spieren. De spiercellen bestaan uit drie delen:
- Een niet samentrekbaar deel met de celkernen, de mitochondria en het Golgi-apparaat.
- Een samentrekbaar deel dat bestaat uit actine- en myosinevezels.
- Een niet samentrekbaar deel van de spiercellen dat verbindingen kan maken met andere spiercellen of zenuwen.
Er zijn drie typen spiercellen:
- Circomyariane spiercel, waarbij de spiervezels het sarcoplasma, het ongedifferentieerde protoplasma van de spiercel, volledig omringen.
- Coelomyariane spiercel, waarbij de spiervezels aangrenzend en loodrecht op de hypodermis zijn en zich langs verschillende kanten langs de spiercel uitstrekken, zoals bij Mermis nigrescens.[40]
- Platymyariane spiercel, waarbij de spiervezels allemaal aan de hypodermis grenzen en er loodrecht op staan, zoals bij Caenorhabditis elegans.[41]
De relatief stijve cuticula vormt samen met de spieren een hydroskelet, omdat rondwormen voor het grootste deel geen spieren in de omtrek hebben. Bij de kop en de anus zitten wel spieren in de omtrek. De inwendige druk van het hydroskelet ligt tussen 70 en 210 mmHg. Samen met de in de lengte liggende spieren als antagonisten kan de rondworm zich als een slang voortbewegen of een deel van zijn lichaam omhoog steken.
Zenuwuitsteeksels lopen vanuit het binnenste oppervlak van de spiercellen naar de aan de buikzijde gelegen zenuwbaan. Dit is uniek in het dierenrijk, waarbij de zenuwcellen uitlopers hebben naar de spieren in plaats van anders om.[35]
Een mannelijke rondworm bestaat van voor naar achteren uit een mondopening, darm (spijsverteringskanaal), cloaca, excretie-orgaan, testikel (mannelijk orgaan) en een excretie-opening. Aan de staartzijde kunnen een of twee spicula, die dienen om de vulva van vrouwelijke rondwormen te openen en de overdracht van sperma te vergemakkelijken, hoewel sperma niet rechtstreeks door of met de spicula wordt overgedragen.[42][43] Het gubernaculum is een ander orgaan van het mannelijk copulatiesysteem, dat de spiculum of de spicula tijdens de copulatie geleidt. Alleen Dichelyne alatae heeft aan de staartzijde ter hoogte van de cloaca-opening korte, brede vleugels, gevleugelde, ongelijke spicula en een schildvormig gubernaculum.
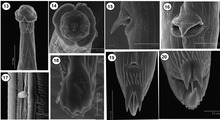 13: Subventraal aanzicht van het voorste uiteinde van Torrestrongylus tetradorsalis met 'kopschermen', 14: Frontaalaanzicht met mondopening, papillen en amfiden.
13: Subventraal aanzicht van het voorste uiteinde van Torrestrongylus tetradorsalis met 'kopschermen', 14: Frontaalaanzicht met mondopening, papillen en amfiden. Mannelijk staarteind van Capillaria plectropomi met de spinose spiculumschede en de uitgeschoven spiculum.
Mannelijk staarteind van Capillaria plectropomi met de spinose spiculumschede en de uitgeschoven spiculum. Staarteind van de mannelijke rondworm Gongylonema pulchrum met rechts het spiculum en links het gubernaculum.
Staarteind van de mannelijke rondworm Gongylonema pulchrum met rechts het spiculum en links het gubernaculum. Tekeningen van een mannelijke Philometra-rondworm - E en F zijn het gubernaculum.
Tekeningen van een mannelijke Philometra-rondworm - E en F zijn het gubernaculum. Links boven: eileider met rijpe eieren, rechts boven: c=vulva en e=vagina, f=baarmoeder, onder:eieren met bissi van Mermis nigrescens
Links boven: eileider met rijpe eieren, rechts boven: c=vulva en e=vagina, f=baarmoeder, onder:eieren met bissi van Mermis nigrescens




Een vrouwelijke rondworm heeft als vrouwelijke organen een eierstok met een eileider, twee spermazakjes, baarmoeder en in het midden van het lichaam een vulva.
Rondwormen hebben een simpel zenuwstelsel dat uit twee banen bestaat. De zenuwbaan aan de rugzijde (dorsaal) is groter dan die aan de buikzijde (ventraal). De strengen worden aan de voorkant om de farynx en soms ook richting het uiteinde met elkaar verbonden door dwarsverbindingen.[44]
De lichaamsholte is een pseudocoeloom waarin vocht zit, dat helpt voedingsstoffen en zuurstof te verspreiden. In de pseudocoeloom kunnen fagocyte witte bloedcellen (coelomocyten) voorkomen. Ze zijn afkomstig van mesenchymatische stamcellen. Deze fagocyte witte bloedcellen zijn variabel van vorm en variëren van ronde, eivormige, kubusvormige en spoelvormige cellen tot stervormige of vertakte cellen die meestal op vaste posities in de pseudocoelom zitten. Ze zijn betrokken bij de afweer tegen ongewenste bacteriën door onder andere het uitscheiden van antibacteriële stoffen.[45]
Hun huid is bedekt met een cuticulum dat tegen uitdrogen beschermt. Hoewel dit cuticulum beweging en beperkte vormveranderingen toestaat groeit het niet mee en rondwormen moeten dan ook als ze groeien regelmatig vervellen. Er zijn vijf larvale stadia, L1, L2, L3 en L4. L5 is het volwassen stadium. In het ei vinden twee vervellingen plaats.
Mond
De mond heeft drie of zes lippen, die vaak een reeks tanden aan hun binnenranden dragen. In de mondholte kan een al of niet holle stilet voorkomen, waarmee de rondworm een cel kan aanprikken om deze vervolgens met de holle stilet leeg te zuigen. De stilet is een geharde, uitstekende opening van de darm (spijsverteringskanaal). Aan de basis van de stilet zitten twee knobbels.[46] Plantenparasitaire aaltjes, zoals Pratylenchus-soorten hebben een stilet. Ook kan er aan de binnenkant van de wang een 'tand'-achtige structuur zitten, waarmee de rondworm een opening in de cuticula van een insect kan maken, zoals bij Heterorhabditis bacteriophora. Op de lippen kunnen setae zitten, stijve, maar toch buigzame, haarvormige structuren.[47][48] De vorm van de mond hangt af van het voedsel. Er worden vijf groepen rondwormen naar hun mondvorm onderscheiden: bacterivoor, fungivoor, plantenparasitaire aaltjes, predator en omnivoor.[49]
Bij Pristionchus-soorten, zoals Pristionchus pacificus treedt dimorfisme in de monddelen op, waardoor deze rondwormen zich op verschillende voedselbronnen kunnen specialiseren. Een type met een kleine mond en een type met een grote mond met tanden.[50] Bij Pristionchus borbonicus komen afhankelijk van het aanwezige voedsel zelfs vijf verschillende mondtypen voor.
 Stilet bij het plantenparasitaire aaltje Pratylenchus sp.
Stilet bij het plantenparasitaire aaltje Pratylenchus sp. De mond van een hondenspoelworm. Afbeelding met een rasterelektronenmicroscoop.
De mond van een hondenspoelworm. Afbeelding met een rasterelektronenmicroscoop. Mondopening met tanden van Parapristionchus giblindavisi
Mondopening met tanden van Parapristionchus giblindavisi
.jpg)


Spijsverteringskanaal

De mondholte is bekleed met de cuticula, die vaak wordt versterkt met structuren, zoals richels, en vooral bij vleesetende soorten een aantal tanden kan hebben. De mondholte mondt uit in een gespierde, zuigende farynx (keelholte), ook bekleed met de cuticula. Spijsverteringsklieren zitten in dit deel van het spijsverteringskanaal, die de enzymen voor de vertering uitscheiden. Bij soorten met een stilet kunnen deze verteringsenzymen in de prooi worden geïnjecteerd.[35] Aan het eind van de farynx zit een verdikking voor het vermalen van het voedsel.

Er is geen maag aanwezig. De farynx sluit aan op de slokdarm (oesofagus), die aansluit op een spierloos gedeelte, de darm, dat de grootste lengte van het spijsverteringskanaal omvat. Hier worden meer enzymen geproduceerd en worden de voedingsstoffen door een één cel dikke laag geabsorbeerd. Het laatste deel van het spijsverteringskanaal is het rectum dat ook bekleed is met de cuticula. Het achterste deel van het rectum tot aan de anus wordt het proctodeum genoemd. Net onder en voor de punt van de staart zit de anus. Door de lichaamsbewegingen van de rondworm wordt het voedsel door het spijsverteringkanaal voortbewogen. Het spijsverteringskanaal heeft kleppen of sluitspieren aan beide uiteinden voor het beheersen van de beweging van het voedsel door het lichaam.[35]
Bij de Trichocephalida en Mermithida is in sommige stadia een zeer opvallend stichosoom aanwezig, die bestaat uit een in de lengterichting liggende reeks van klierachtige, eencellige cellen, stichocyten, gerangschikt in een rij langs de farynx en de achterste farynxklieren vormen. Het mondt uit in de farynxholte en functioneert blijkbaar als een uitscheidingsklier en opslagorgaan. De stichocyten bevatten mitochondria, ruw endoplasmatisch reticulum, veel Golgi-apparaten en meestal een van twee soorten uitgescheiden granulen, α-granulen en β-granulen, die duiden op de uitscheidingsfunctie van de stychocyt.
De spijsvertering kan uitwendig zijn, waarbij de rondworm verteringsenzymen in de prooi brengt of inwendig, waarbij de prooi wordt ingeslikt. Zo brengt de dennenhoutnematode (Bursaphelenchus xylophilus) cellulasen in de prooi. In het spijsverteringskanaal kunnen ook endosymbiotische micro-organismen (bacteriën of schimmels) voorkomen, die nodig zijn voor de afbraak van bepaalde voedingsbestanddelen zoals cellulose.
Staart
Aan het uiteinde van de staart wordt vaak een kleverige 'staartklier' gevonden.[35] De uitgangsopening van de klier wordt een spinneret genoemd, zoals bij Dichromadora geophila. De staart kan veel verschillende vormen hebben. Draadvormig, geleidelijk versmallend, kegelvormig, vingervormig, enigszins vingervormig, bol-kegelvormig met een korte punt, bol-kegelvormig met een lange punt, kegelvormig met een gekromde punt, kegelvormig met een stompe punt, kegelvormig met naar de buikzijde boogvormige punt, rond-cilindervormig, stekelpuntig, meervoudig-vingervormig.[51]
Uitscheidingssysteem
Het teveel aan stikstof wordt in de vorm van ammoniak via de lichaamswand uitgescheiden en is niet gebonden aan specifieke organen. De structuren voor het uitscheiden van zout voor het op peil houden van de osmoregulatie zijn echter meestal complexer.[35]
Bij veel in zee levende rondwormen scheiden een of twee eencellige 'renette' klieren het zout uit in de vorm van 0,5-1,5 µm grote korrels door een porie aan de onderkant van het lichaam, even voorbij de farynx. Bij de meeste andere rondwormen zijn deze gespecialiseerde cellen vervangen door een orgaan bestaande uit twee parallelle kanaaltjes verbonden door een enkel dwars kanaaltje. Dit dwarse kanaaltje komt uit in een gemeenschappelijk kanaaltje dat uitkomt bij de uitscheidingsopening.[35] Met de uitscheidingen kunnen de rondwormen zich vastzetten op de ondergrond.[52]
Zenuwstelsel

Vier perifere zenuwen lopen over de lengte van het lichaam, dorsaal (rugzijde), ventraal (buikzijde) en lateraal (zijkant). Elke zenuw wordt omgeven door bindweefsel dat onder de huid en tussen de spiercellen ligt. De ventrale zenuw is de grootste en heeft een dubbele structuur naar de excretieopening. De dorsale zenuw is verantwoordelijk voor motorische controle, terwijl de laterale zenuwen sensorisch van aard zijn en de ventrale zenuw combineert beide functies.[35]
De enige plaats op het lichaam van de rondworm waar onbeweeglijke trilharen voorkomen, en die een sensorische functie hebben, zijn de zenuwbanen.[53][54]
Aan het voorste uiteinde van de rondworm vertakken de zenuwen zich vanuit een dichte, cirkelvormige zenuw (zenuwring) rondom de farynx en dient als een soort hersenen. Kleinere zenuwen lopen naar voren vanuit de ring voor de sensorische organen van de kop.[35]
De lichamen van de rondwormen zijn bedekt met talloze sensorische haren en papillen die samen zorgen voor een gevoel van aanraking. Achter de sensorische haren op de kop liggen twee kleine putjes, of amfiden. Dit zijn de belangrijkste reukorganen van een rondworm. Elke amfide bestaat uit twaalf sensorische zenuwcellen en met trilhaartjes bezette dendrieten. Enkele in het water levende rondwormen bezitten wat pigmentvlekken, die op oogvlekken lijken, maar het is onduidelijk of deze daadwerkelijk sensorisch van aard zijn.[35]
"Spierarmen" zijn in de huid voorkomende armvormige spiermembraanverlengingen, die de in de huid voorkomende spiermembranen met de motoraxonen van de dorsale zenuwbaan en die van de ventrale zenuwbaan verbinden. In tegenstelling tot andere organismen waar neuronen prikkels naar hun doelspiercellen sturen om synapsen te maken, worden neuromusculaire knooppunten van Caenorhabditis elegans gemaakt door armvormige uitgroeiingen (spierarmen) van de spiercellen naar de motorneuronen. Deze spierarmen bestaan uit eenvoudige structuren en hebben een steel met een vertakte terminus die contact maakt met het neuron. Vergelijkbaar met chemische synapsen tussen neuronen, worden neuromusculaire knooppunten terloops gemaakt door de gestimuleerde neuronen aan deze spierarmen.[55]
Ei

.jpg)
Eieren kunnen een embryo of nog geen embryo bevatten op het moment dat ze door het vrouwtje worden uitgescheiden. Er is nog geen embryo aanwezig als de bevruchte eieren nog niet zijn ontwikkeld. Van een paar soorten is bekend dat ze eierlevendbarend zijn. Het omhulsel van het ei bestaat uit drie lagen, met als buitenste laag de vitellinelaag, dan een chitine-achtige middelste laag en een binnenste laag bestaande uit lipiden. De vitellinelaag bestaat voornamelijk uit eiwitvezels, met eiwitreceptoren die nodig zijn voor de spermabinding die op hun beurt gebonden zijn aan spermaplasmamembraanreceptoren. Bij vrij levende rondwormen vinden de eerste en tweede larvale stadia al in het ei plaats. Bij parasitaire rondwormen is de levenscyclus vaak veel gecompliceerder.[35]
Sommige rondwormsoorten, zoals Mermis nigrescens hebben eieren met aan de polen vertakte filamenten (byssus, meervoud byssi).[56] Deze filamenten zijn vertakkingen van het eiwitmembraan van het ei en zorgen ervoor dat het ei zich kan vasthechten aan de ondergrond.
 Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans met stralen (sensory rays) in de bursa (In werkelijkheid is het aaltje kleurloos en doorschijnend.)
Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans met stralen (sensory rays) in de bursa (In werkelijkheid is het aaltje kleurloos en doorschijnend.) Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans met stralen (sensory rays) in de bursa (In werkelijkheid is het aaltje kleurloos en doorschijnend.)
Lichaamsbouw van de mannelijke organen van een Caenorhabditis elegans met stralen (sensory rays) in de bursa (In werkelijkheid is het aaltje kleurloos en doorschijnend.) Lichaamsbouw van de vrouwelijke organen van de hermafrodiete Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.)
Lichaamsbouw van de vrouwelijke organen van de hermafrodiete Caenorhabditis elegans (In werkelijkheid is het aaltje kleurloos en doorschijnend.) Trilobus longus. I, mannetje; II, kop, zijaanzicht; III, kop, zijaanzicht; IV, kop, buikaanzicht; V, achterste supplementair orgaan; VI, achterste supplementair orgaan; VII, twee supplementaire organen van een buitengewoon vrouwtje. a, seta aan zijkant; b, papillen; c, seta iets naast het midden; d, farynx; e, orgaan aan zijkant; f, tand; g, tand; h, slokdarm (oesofagus); i, zenuwring; j, uitscheidingsopening; k, lichaamsspieren; l, klieren; m, spijsverteringskanaal (darm); n, dichteind voorste testikel; 0, testikel; p, verbindingsplaats testikels; 5, dichteind achterste testikel; r, zaadleider: s, zenuw van supplementair orgaan; t, holte van supplementair orgaan; u, linker spiculum; v, gubernaculum; w, de drie staartklieren; x, anus; y, staartpunt; z, top van supplementair orgaan. (Naar Cobb.)
Trilobus longus. I, mannetje; II, kop, zijaanzicht; III, kop, zijaanzicht; IV, kop, buikaanzicht; V, achterste supplementair orgaan; VI, achterste supplementair orgaan; VII, twee supplementaire organen van een buitengewoon vrouwtje. a, seta aan zijkant; b, papillen; c, seta iets naast het midden; d, farynx; e, orgaan aan zijkant; f, tand; g, tand; h, slokdarm (oesofagus); i, zenuwring; j, uitscheidingsopening; k, lichaamsspieren; l, klieren; m, spijsverteringskanaal (darm); n, dichteind voorste testikel; 0, testikel; p, verbindingsplaats testikels; 5, dichteind achterste testikel; r, zaadleider: s, zenuw van supplementair orgaan; t, holte van supplementair orgaan; u, linker spiculum; v, gubernaculum; w, de drie staartklieren; x, anus; y, staartpunt; z, top van supplementair orgaan. (Naar Cobb.)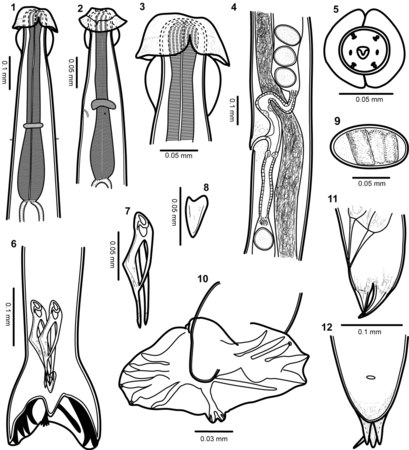 Lichaamsbouw van Torrestrongylus tetradorsalis. 1: Voorste uiteinde van mannetje, buikaanzicht, met de farynx en de relatieve posities van de zenuwring en deiriden (sensorische papillen als tastorgaan in het laterale cervicale gebied); let op scheiding van cuticulaire projectie in voorste blaasje. 2: Voorste uiteinde van vrouwtje, zijaanzicht, dat de relatieve posities van de zenuwring, deiride en uitscheidingsporie toont; let op continue cuticulaire expansie in voorste blaasje. 3: Detail van de voorste helft van het kopblaasje met de laterale uitsteeksels die een paraplu vormen en de achterste helft met de vorm van de "twee handvatten van een kruik". 4: Ovejector (het terminale, zeer gespierde deel van de eileider, die het ei door de genitale porie duwt) van vrouwtje met flap in voorste lip van vulva, vagina vera, infundibulum (tweede deel van de eileider), sluitspieren en baarmoedervertakkingen met uitgegroeide eieren. 5: Gezichtsaanzicht van het mannetje, met de rangschikking van papillen en cuticulaire vergroting van de voorste kopblaasje. 6: Achterste uiteinde van het mannetje, dat de relatieve positie van spicula en gubernaculum toont. 7: Spiculum. 8: Gubernaculum. 9: Ei. 10: Bursa met plaats en vertakking van dorsale straal. 11: Zijaanzicht van de staart van het vrouwtje. 12: Buikaanzicht van vrouwelijke staart.
Lichaamsbouw van Torrestrongylus tetradorsalis. 1: Voorste uiteinde van mannetje, buikaanzicht, met de farynx en de relatieve posities van de zenuwring en deiriden (sensorische papillen als tastorgaan in het laterale cervicale gebied); let op scheiding van cuticulaire projectie in voorste blaasje. 2: Voorste uiteinde van vrouwtje, zijaanzicht, dat de relatieve posities van de zenuwring, deiride en uitscheidingsporie toont; let op continue cuticulaire expansie in voorste blaasje. 3: Detail van de voorste helft van het kopblaasje met de laterale uitsteeksels die een paraplu vormen en de achterste helft met de vorm van de "twee handvatten van een kruik". 4: Ovejector (het terminale, zeer gespierde deel van de eileider, die het ei door de genitale porie duwt) van vrouwtje met flap in voorste lip van vulva, vagina vera, infundibulum (tweede deel van de eileider), sluitspieren en baarmoedervertakkingen met uitgegroeide eieren. 5: Gezichtsaanzicht van het mannetje, met de rangschikking van papillen en cuticulaire vergroting van de voorste kopblaasje. 6: Achterste uiteinde van het mannetje, dat de relatieve positie van spicula en gubernaculum toont. 7: Spiculum. 8: Gubernaculum. 9: Ei. 10: Bursa met plaats en vertakking van dorsale straal. 11: Zijaanzicht van de staart van het vrouwtje. 12: Buikaanzicht van vrouwelijke staart. Lichaamsbouw van Capillaria plectropomi. A: voorste uiteinde van mannetje, zijaanzicht. B: stichocyte in het middelste deel van het stichosoom. C: achterste uiteinde van mannetje, zijaanzicht. D: kop van vrouwtje, apicaalaanzicht. E: staarteinde van mannetje, ventraalaanzicht. F: regio van vulva, zijaanzicht. G, H: staarteinde van mannetje (verschillende exemplaren), zijaanzichten. I: staart van vrouwtje, zijaanzicht. J: spiculum, zijaanzicht. K: laterale bacillaire band in de farynx, zijaanzicht. L: volledig ontwikkeld ei.
Lichaamsbouw van Capillaria plectropomi. A: voorste uiteinde van mannetje, zijaanzicht. B: stichocyte in het middelste deel van het stichosoom. C: achterste uiteinde van mannetje, zijaanzicht. D: kop van vrouwtje, apicaalaanzicht. E: staarteinde van mannetje, ventraalaanzicht. F: regio van vulva, zijaanzicht. G, H: staarteinde van mannetje (verschillende exemplaren), zijaanzichten. I: staart van vrouwtje, zijaanzicht. J: spiculum, zijaanzicht. K: laterale bacillaire band in de farynx, zijaanzicht. L: volledig ontwikkeld ei.



_(20789723172).jpg)


Levenscyclus
Samenvatten
Perspectief
.jpg)


Als voorbeeld wordt die van de wortelknobbelaaltjes beschreven.

_penetrates_a_tomato_root_-_USDA-ARS.jpg)

_in_Arachis_hypogaea.jpg)
Alle rondwormen hebben een embryonaalstadium, vier larvale stadia (L1 - L4) en een volwassen stadium. Het jonge Meloidogynes-aaltje komt uit het ei als een vermiforme larve in het tweede stadium (L2), waarbij de eerste vervelling in het ei heeft plaatsgevonden. Pas uitgekomen larven hebben een korte vrijlevende fase in de bodem, in de rhizosfeer van de waardplanten. Ze kunnen de ouderwaardplanten opnieuw infecteren of door de grond migreren naar een nieuwe waardplantwortel. L2-larven voeden zich niet tijdens de vrijlevende fase, maar leven van in de darm opgeslagen lipiden.[57]
Een uitstekend modelsysteem voor de studie van het parasitaire gedrag van plantparasitaire aaltjes is ontwikkeld met behulp van de zandraket (Arabidopsis thaliana) als modelgastheer.[58] De wortels van de zandraket zijn aanvankelijk klein en transparant, zodat elk detail zichtbaar is. Binnendringen en migratie in de wortel werd bestudeerd met Meloidogyne incognita.[59] Aaltjes van het tweede larvale stadium dringen het wortelstrekkingsgebied binnen en migreren in de wortel totdat ze een vaste verblijfplaats hebben gevonden. Signalen van de L2-aaltjes zorgen ervoor dat parenchymcellen in de buurt van de kop van het L2-aaltje meerkernig worden[60] voor het vormen van voedingscellen, algemeen bekend als reuzencellen, van waaruit de L2 en later de volwassen aaltjes zich voeden met behulp van een stilet.[61] Gelijktijdig met de vorming van reuzencellen, vormt het omringende wortelweefsel een gal (knobbel) waarin de zich ontwikkelende larve zit. De larven beginnen zich met de inhoud van de reuzencellen te voeden ongeveer 24 uur nadat ze een vaste verblijfplaats hebben gevonden.
Na verdere voeding ondergaan de L2-aaltjes morfologische veranderingen en worden ze bolvormig. Zonder verdere voeding vervellen ze drie keer en worden uiteindelijk volwassen. Bij vrouwtjes, die bijna bolvormig zijn, wordt de voeding hervat en ontwikkelt het voortplantingssysteem zich.[57] De mannetjes kunnen zich vrij bewegen en verlaten de wortel nadat ze volwassen zijn geworden. De levensduur van een volwassen vrouwtje kan tot drie maanden oplopen en er kunnen honderden eieren worden geproduceerd. Vrouwtjes kunnen na de oogst van de bovengrondse delen van de plant doorgaan met het leggen van eieren. De overlevingsfase van het aaltje tussen de opeenvolgende gewassen bevindt zich meestal in het ei.
De lengte van de levenscyclus is temperatuurafhankelijk.[62][63] De relatie tussen ontwikkelingssnelheid en temperatuur is lineair over een groot deel van de levenscyclus van de wortelknobbelaaltjes, hoewel het mogelijk is dat de verschillende stadia van de levenscyclus, bijvoorbeeld ei-ontwikkeling, gastheer wortelinvasie van de waardplant of groei, een iets ander optimum hebben. Soorten binnen het geslacht Meloidogyne kunnen ook verschillende temperatuuroptima's hebben. Bij bijvoorbeeld Meloidogyne javanica vindt de ontwikkeling plaats tussen 13 en 34 °C, met een optimale ontwikkeling bij ongeveer 29 °C.
De vrouwtjes leggen de eieren in een gelatineuze massa geproduceerd door zes rectale klieren en afgescheiden voor en tijdens het leggen van de eieren.[64][65] De gelatineuze massa vormt aanvankelijk een kanaal door de buitenste lagen van het wortelweefsel en omringt later de eieren. De gelatineuze massa vormt een barrière tegen waterverlies en zorgt door het hoge vochtgehalte dat de eieren niet uitdrogen.[66] Naarmate de gelatineuze massa ouder wordt, verandert het van een kleverige, kleurloze gelei in een leerachtige, oranje-bruine substantie die gelaagd lijkt.[67]
De eivorming is in Meloidogyne javanica gedetailleerd bestudeerd[68] en lijkt op de eivorming in het goed bestudeerde, vrij levende aaltje Caenorhabditis elegans.[69] Ook de embryogenese is bestudeerd en de ontwikkelingsstadia in een geplette eimassa zijn gemakkelijk te identificeren met een fasecontrastmicroscoop. Het ei bestaat uit één cel, met herkenbare volgende stadia van twee cellen, vier cellen en acht cellen. Uiteindelijk is de morula gevormd. Verdere celdeling leidt tot het kikkervisjesstadium, waarbij door een verdere verlenging het eerste larvale stadium ontstaat. Het aaltje is nu ruwweg vier keer zo lang als het ei was. Het L1-stadium van Caenorhabditis elegans heeft 558 cellen en het L1-stadium van Meloidogyne javanica heeft waarschijnlijk een vergelijkbaar aantal, omdat alle aaltjes morfologisch en anatomisch vergelijkbaar zijn.[69] Het omhulsel van het ei bestaat uit drie lagen, met als buitenste laag de vitellinelaag, dan een chitine-achtige middelste laag en een binnenste laag bestaande uit lipiden. De vitellinelaag bestaat voornamelijk uit eiwitvezels, met eiwitreceptoren die nodig zijn voor de spermabinding die op hun beurt gebonden zijn aan spermaplasmamembraanreceptoren.
Nadat het eiomhulsel doorlaatbaar is geworden, kan de larve uit het ei komen met behulp van fysieke of enzymatische processen.[70] Eitjes van de wortelknobbelaaltjes worden in het algemeen niet beïnvloed door de aanwezigheid van een gastheer, maar komen pas uit bij de juiste temperatuur en in aanwezigheid van water. Ammonium-ionen kunnen het uitkomen remmen en het binnendringingsvermogen in de plantenwortels door de Meloidogyne incognita -larven verminderen.[71]
Dauerlarve
Van sommige rondwormsoorten kan de larve in rust gaan, in een soort stase-toestand. Zo'n larve wordt een dauerlarve genoemd en kan in deze toestand onder ongunstige omstandigheden tot wel vier maanden overleven. Met name komt dit voor bij Rhabditidae-larven, waaronder Caenorhabditis elegans-larven. Omdat het moment van overgaan in het dauerstadium afhankelijk is van omgevingsfactoren, vertegenwoordigt het een klassiek en goed bestudeerd voorbeeld van polyfenisme.[72][73]

In 1975 beschreven Cassada en Russell het larvale dauerstadium van Caenorhabditis elegans.[74] Bij C. elegans gaat aan het einde van het eerste larvale stadium (L1) de larve over in een dauerlarve onder invloed van populatiedichtheid, gebrek aan voedsel en temperatuurverhoging, waarbij de populatiedichtheid de grootste invloed heeft. Door een bepaald feromoon wordt de hele populatie aangezet om in het dauerstadium te gaan. Een hydrofiele thermohardende stof remt de overgang naar het dauerstadium en haalt de larve weer uit het dauerstadium.
Tijdens het dauerstadium is de larve min of meer plat en is de 'huid' taaier dan bij een niet in rust verkerende larve. In deze toestand is de mondholte door een soort binnenkap afgesloten en is er geen peristaltiek. Dauerlarven hebben hoge niveaus van het mRNA dat codeert voor het hitteschokeiwit Hsp90, het superoxidedismutase en katalase. C. elegans en Pristionchus pacificus hebben het DAF-12-endocriene systeem voor de vorming van dauerlarven.
In het dauerstadium zijn bij vele soorten 'alae’ zichtbaar, een of twee ribbels gevormd door onderhuids weefsel.[75] Ze komen ook voor bij volwassen rondwormen.
Het dauerstadium heeft andere namen bij de verschillende soorten rondwormen zoals 'diapauze' of 'hypobiose', maar omdat de rondworm C. elegans de meest bestudeerde rondworm is geworden, wordt de term 'dauerstadium' of 'dauerlarve' algemeen erkend, wanneer naar deze toestand wordt verwezen in andere vrijlevende aaltjes.
Het dauerstadium wordt ook beschouwd als gelijkwaardig aan het infectieuze stadium van parasitaire rondwormlarven.
Anhydrobiotische rusttoestand
Sommige rondwormen, zoals het aardbeibladaaltje, kunnen in een anhydrobiotische rusttoestand langere tijd overleven. Door het verlies van hun lichaamsvocht verschrompelen ze, en bovendien maken ze trehalose aan, een suiker die voorkomt dat hun weefsels door de uitdroging beschadigd raken.
L1-cyste
Trichinella-soorten vormen L1-cysten in het sperweefsel van hun gastheer. Deze L1-cysten kunnen tot 30 jaar infectieus blijven. De L1-larven gaan nadat ze door het vrouwtje uitgescheiden zijn via de haarvaten in de dwarsgestreepte spieren naar de spiervezelcellen en vormen daar voedingscellen. In slechts drie weken veroorzaken de larven grote veranderingen in deze spiercellen. Ze vergroten onder andere de celkern, veroorzaken kerndelingen en creëren een "placenta"-achtige structuur rond de spiercel, een bloedvatennetwerk. De vorming van nieuwe bloedvaten rond de spiercel wordt waarschijnlijk geïnduceerd doordat genen van de larven bepaalde genen van de cel van de gastheer activeren.
Levend barend
Bij sommige soorten rondwormen, zoals Heterorhabditis bacteriophora, Steinernema glaseri en Steinernema carpocapsae, kunnen de larven zich in het begin ontwikkelen in de uterus. Op een gegeven moment verlaten ze die en voeden de ontwikkelende larven zich eerst met de bacteriën in het spijsverteringskanaal en later met de weefsels van de dode moeder. De larven van het geslacht Steinernema verlaten het moederlichaam via natuurlijke openingen, terwijl die van het geslacht Heterorhabditis door de taaie 'huid' breken.[76] Dit wordt endotokia matricida genoemd.[77] Ook bij de rode lebmaagworm (Haemonchus contortus), Mehdinema allii en Metacrobeles amblyurus treedt dit verschijnsel op.[78] Vooral onder stressomstandigheden kan dit verschijnsel optreden. Ook Trichinella spiralis is levendbarend, waarbij de L1-larven via de anus het moederlichaam verlaten en in het spierweefsel cysten vormen.
Microfilariae

Bij de tropische ziekten filariasis en rivierblindheid worden door de parasitaire rondworm eerst microfilariae gevormd, die later overgaan in L1-larven. De oude eischaal zit nog om de microfilaria heen en wordt pas in de steekmug afgebroken.
De parasitaire rondwormen uit de orde Spirurida, die filariasis beroorzaken, worden door muggen verspreid. De rondworm leeft in de lymfevaten van de mens, waarbij door de rondworm in zijn leven miljoenen microfilariae worden geproduceerd. Deze microfilariae circuleren in de bloedstroom, vooral rond middernacht. Als een mug de patiënt steekt komen de microfilariae in het spijsverteringskanaal van de mug. In de mug verplaatsen ze zich naar de borstspieren om in enkele weken tijd uit te groeien tot infectieuze L1-larven. Deze migreren naar de kop en de zuigsnuit van de mug, zodat ze bij een muggenbeet opnieuw iemand kunnen infecteren. Het gaat vooral om de rondwormen Wuchereria bancrofti en Brugia malayi. Eenmaal door de huid begeven ze zich naar de lymfevaten waar ze uitgroeien tot volwassen rondwormen.
Bij rivierblindheid gaat het om een infectie met Onchocerca volvulus die in Afrika meestal wordt verspreid door kriebelmuggen (Simulium damnosum), in een aantal Afrikaanse landen door andere soorten kriebelmuggen zoals Simulium neavei. Deze kriebelmuggen houden zich graag op in de buurt van snelstromende rivieren en bijten overdag. Na een beet van de kriebelmug gaan de larven van deze rondwormen met het speeksel van de mug mee naar de bloedbaan waarna de larven zich naar een plaats vlak onder de huid begeven waar ze knobbeltjes vormen. Op deze plaats vindt de geslachtelijke voortplanting plaats. De vrouwelijke rondworm produceert dagelijks tot wel enkele duizenden eieren die rijpen tot microfilariae.
Wijze van voortplanting
Samenvatten
Perspectief



Rondwormen kunnen eenslachtig of hermafrodiet zijn. De meeste rondwormen zijn eenslachtig met zowel vrouwelijke als mannelijke exemplaren. Bij sommige soorten zoals Caenorhabditis elegans zijn de meeste rondwormen hermafrodiet en komen er slechts enkele mannelijke exemplaren voor. Rondwormsoorten vertonen een reeks van verschillende wijzen van voortplanting, waaronder seksuele voortplanting (amfimixis), facultatieve seksuele voortplanting, meiotische parthenogenese (automixie) en mitotische parthenogenese (apomixie).[80] De wijze van voortplanting kan eenvoudig tot ingewikkeld zijn. Vooral parasitaire soorten hebben soms een ingewikkeld levens- en voortplantingscyclus met een generatiewisseling tussen gastheren of organen. Zo heeft Onchocerca volvulus een gastheerwisseling met de mens en de kriebelmug en heeft Strongyloides stercoralis een ingewikkelde wijze van facultatieve seksuele voortplanting.
Beide geslachten bezitten een of twee buisvormige gonaden. Bij de mannetjes wordt het sperma geproduceerd aan het einde van de geslachtsklieren en verplaatst het zich tijdens de rijping over de lengte van de rondworm. De testis komt uit in een relatief brede zaadblaas en gaat vervolgens over in een beklierd en gespierd ejaculatiekanaal dat te vergelijken is met de zaadleider en cloaca. Bij de vrouwtjes komen de twee eierstokken uit in twee eileiders (bij hermafrodieten komen de eieren eerst in een spermatheca) en vervolgens in een beklierde baarmoeder. De baarmoeders komen beide uit in een gemeenschappelijke vulva/vagina, meestal gelegen aan de buitenkant in het midden van de morfologisch buikzijde.[35]
De voortplanting is meestal seksueel, hoewel hermafrodieten in staat zijn tot zelfbevruchting. Mannetjes zijn meestal kleiner (vaak veel kleiner) dan vrouwtjes of hermafrodieten en hebben vaak een karakteristiek gebogen of waaiervormige staart. Aan de staartzijde van de mannelijke rondworm kunnen een of twee spicula voorkomen, die dienen om de vulva van de vrouwelijke rondwormen te openen en de overdracht van sperma te vergemakkelijken, hoewel sperma niet rechtstreeks door of met de spicula wordt overgedragen.[42][43] Het gubernaculum is een ander orgaan van het mannelijk copulatiesysteem, dat de spiculum of de spicula tijdens de copulatie geleidt. Tijdens de copulatie kruipen amoebeachtig zaadcellen langs het spiculum in de vulva van het vrouwtje. Aangenomen wordt dat de zaadcel van rondwormen de enige eukaryotische cel is zonder het bolvormige eiwit G-actine. Zogenaamde supplementaire organen kunnen voorkomen bij zowel mannetjes als vrouwtjes. Het zijn uitscheidings- en aanhechtingsorganen, die bij de paring een rol spelen. (Zie tekening Tobrilus longus. Ze worden door de cuticula gevormd.
Bij Trichosomoides crassicauda zitten de mannelijke rondwormen in de baarmoeder of vagina van de volwassen vrouwelijke rondworm.[81]
Bijzondere vormen van voortplanting
Sommige hermafrodiete rondwormen, zoals Heterorhabditis-soorten van de orde Rhabditida, hebben een wijze van voortplanting, genaamd endotokia matricida (in de uterus geboren), wat de dood van de moeder tot gevolg heeft. Dit komt vooral voor bij entomopathogene rondwormen, rondwormen die van insecten leven. De larven eten de moeder van binnen uit op en laten alleen het exoskelet achter. Deze wijze van voortplanting verzekert de ontwikkeling van de larven op een moment dat de externe voedselvoorziening afneemt en zorgt ervoor dat de nakomelingen voldoende energiereserves en symbiotische bacteriën hebben, waardoor ze voor lange tijd zonder voedsel kunnen overleven.[82][83]
Caenorhabditis elegans en Caenorhabditis briggsae hebben androdioecie als voortplantingmethode, wat bij dieren zeer zeldzaam is. Hierbij komen zowel mannetjes als hermafrodieten tegelijk voor. De hermafrodieten geven echter bij de voortplanting de voorkeur aan de mannetjes in plaats van aan zelfbevruchting te doen. Ook geeft het paren met mannetjes meer nageslacht, doordat er dan meer eieren gelegd worden.
Rondwormsoorten van het geslacht Meloidogyne (wortelknobbelaaltjes) hebben een aantal verschillende voortplantingsmethoden, waaronder seksuele voortplanting (amfimixis), facultatieve seksuele voortplanting, meiotische parthenogenese (automixie) en mitotische parthenogenese (apomixie).[80]
Het geslacht Mesorhabditis vertoont een ongebruikelijke vorm van parthenogenese, waarbij sperma producerende mannetjes copuleren met vrouwtjes, maar de spermacel niet fuseert met de eicel. Contact met de spermacel is essentieel voor de eicel om met delen te beginnen, maar omdat er geen fusie van de cellen optreedt, draagt het mannetje geen genetisch materiaal over aan de nakomelingen, die in wezen klonen van het vrouwtje zijn.[35]
Taxonomie
Over de systematiek van de rondwormen (nematoden) bestaat geen consensus.
Pathotypen
Bij soorten van plantenparsitaire aaltjes kunnen pathotypen voorkomen. Een pathotype is een systematische eenheid binnen een aaltjessoort, die niet is te onderscheiden op morfologische kenmerken. Pathotypen verschillen van elkaar doordat ze bepaalde resistenties bij plantenrassen van een of enkele waardplantsoorten hebben doorbroken. Zo zijn van Globodera rostochiensis (het aardappelcystenaaltje) momenteel vijf pathotypen bekend: Ro1, Ro2, Ro3, Ro4 en Ro5. Ro1 komt in Nederland het meeste voor en Ro4 en Ro5 het minste. Zo heeft het wortelknobbelaaltje Meloidogyne arenaria drie pathotypen, waarvan pathotype een het meest voorkomt in Amerika op pinda.
Vrij levende soorten
Vrij levende soorten voeden zich met algen, schimmels, kleine dieren, uitwerpselen, dode organismen en levend weefsel. Vrij levende mariene rondwormen zijn belangrijk en komen veel voor bij het meiobenthos. Ze spelen een belangrijke rol in het afbraakproces en helpen bij het opnieuw beschikbaar komen van voedingsstoffen in mariene omgevingen. Ze zijn echter gevoelig voor veranderingen in het milieu veroorzaakt door vervuiling. Caenorhabditis elegans is een in de bodem vrijlevende rondworm, die is gebruikt als modelorganisme. Onder de vrijlevende rondwormen komen plantenparasitaire soorten voor, zoals bladaaltjes, stengelaaltjes en wortellesieaaltjes. Van de wortelknobbelaaltjes zijn de volwassen mannetjes ook vrijlevend.
Parasitaire soorten
Samenvatten
Perspectief




Parasitaire soorten rondwormen, die bij mensen voorkomen, zijn onder andere de Ascaris-soorten, zoals de spoelworm, Filarioidea, Ancylostoma, Necator, aarsmade en zweepworm (Trichuris trichiura). Trichinella spiralis komt voor bij ratten, varkens, beren en bij mensen door het eten van besmet vlees en veroorzaakt de ziekte trichinose of trichinellose. Baylisascaris infecteert meestal wilde dieren, maar kan voor mensen ook dodelijk zijn. De hartworm (Dirofilaria immitis) infecteert de harten, slagaders en longen van honden en sommige katten. De rode lebmaagworm (Haemonchus contortus) is een van de meest voorkomende besmettelijke ziekten over de hele wereld bij schapen.
Entomopathogene rondwormen parasiteren daarentegen op schadelijke insecten en worden door de mens meestal als gunstig beschouwd, maar ze infecteren in sommige gevallen ook nuttige insecten. Wanneer een infectieuze larve een geschikte gastheer heeft gevonden zwelt zijn kop op, opent zijn mond en stulpt zijn spijsverteringskanaal uit.[84] De infectieuze larven dringen de lichaamsholte van de insectenlarve of het insect binnen, meestal via natuurlijke lichaamsopeningen (mond, anus, ademopeningen) of via gebieden met een dunne cuticula. Vervolgens brengen de infectieuze larven mutualistische bacteriën uit hun spijsverteringskanaal in het lichaam van de insectenlarve of het insect, die zich snel in de hemolymfe gaan vermenigvuldigen. De bacteriën doden de gastheer binnen 24-48 uur. Uit onderzoek van 2017 blijkt echter dat Steinernema carpocapsae niet alleen op de bacteriën vertrouwt voor het doden van de gastheer, maar door het uit- en afscheiden van eiwitten meehelpt de gastheer te doden.[85] Samen voeden de rondwormen en bacteriën zich met de door proteasen vloeibaar gemaakte gastheer. De belangrijkste entomopathogene rondwormen, die gebruikt worden bij de biologische bestrijding van schadelijke insecten, behoren tot de geslachten Heterorhabditis en Steinernema.
Een soort rondworm parasiteert alleen de gewone vijgenwesp, terwijl deze wesp het enige insect is die zorgt voor de bevruchting van de vijgen. De bevruchting van de vijgenboom geschiedt namelijk door symbiose met de gewone vijgenwesp, die zowel zichzelf als de vijg voortplant. Een bevruchte vrouwelijke, geparasiteerde wesp gaat de vijg door een opening (ostiole) binnen, bevrucht de vrouwelijke vijgenbloemen met stuifmeel en legt haar eieren in deze bloemen, waarna ze sterft. De nakomelingen van de rondworm gaan met de volgende generatie wespen mee naar nieuwe vijgen.
_&_Mermithid_nematode_(black_background).JPG)
De nieuw ontdekte parasitaire Tetradonematidae-rondworm Myrmeconema neotropicum induceert fruitmimicry bij de tropische mier Cephalotes atratus. Geïnfecteerde mieren ontwikkelen een felrood achterlijf, hebben de neiging om zich trager te bewegen en lopen met hun achterlijf in een opvallend omhoog gerichte positie. Deze veranderingen trekken waarschijnlijk bes etende vogels aan, die de geïnfecteerde mieren opeten. De parasieteneieren komen in de uitwerpselen van de vogels terecht, dat door de mieren worden verzameld om als voedsel voor hun larven te dienen, waarmee de levenscyclus van de rondworm Myrmeconema neotropicum is voltooid.
Evenzo zijn meerdere soorten rondwormen gevonden in de buikholten van de zweetbij, Lasioglossum zephyrum. In het vrouwelijk lichaam belemmert de rondworm de ontwikkeling van de eierstokken en maakt de bij minder actief, dus minder effectief bij het verzamelen van stuifmeel.[86] Ook wordt een Mermithidae-rondworm aangetroffen bij de Aziatische hoornaar.
Bij insecten behorend tot de superfamilie Apoidea werden de volgende rondwormen gevonden. De Dufourklieren van de vrouwtjes van Halictus ligatus, Augochlora pura mosieri en Augochlorella gratiosa (Halictidae) uit Florida waren geïnfecteerd met de rondwormlarven van Acrostichus halicti (Diplogastridae). De Dufourklieren van Colletes thoracicus (Colletidae) vrouwtjes uit Maryland waren geïnfecteerd met de larven van een nieuw soort rondworm behorend tot Koerneria (Diplogastridae) en de klieren in de buikholte van de vrouwelijke Andrena alleghaniensis (Andrenidae) uit New York waren geïnfecteerd met de larven van een andere Koerneria-soort. De eileiders aan de buikzijde en op het mediane vlak, de Dufourklieren en de gifklieren van Anthophora abrupta (Apidae) uit Maryland en Alabama waren geïnfecteerd met de larven van een nieuwe rondwormsoort behorend tot Bursaphelenchus (Aphelenchoididae).[87]
Infectie door parasitaire soorten

De infectie van zoogdieren, inclusief de mens, die dienen als eindgastheer voor de rondworm kan bijvoorbeeld plaatsvinden door:
- het eten van rauw vlees dat besmet is met rondwormlarven van bijvoorbeeld de Trichinella-soort Trichinella spiralis
- het eten van uitwerpselen met rondwormeieren door bijvoorbeeld honden.
- met ontlasting bevattende wormeneieren besmet voedsel als gevolg van slechte hygiëne (bemesting van gewassen met mest, geen handen wassen na wc-bezoek) op te eten.
- dat bij verschillende rondwormsoorten de infectie te wijten is aan actieve penetratie van (filariforme) larven door de huid, bijvoorbeeld de mijnworm of Necator americanus.
- insectenbeten. Bijvoorbeeld Onchocerca volvulus, de veroorzaker van rivierblindheid, wordt meestal verspreid door insectenbeten van kriebelmuggen.
Land-, tuin- en bosbouw




Afhankelijk van de soort kan een rondworm nuttig of schadelijk zijn voor planten. Verschillende planten parasitaire aaltjessoorten, zoals het maïswortelknobbelaaltje en het bedrieglijk maïswortelknobbelaaltje, behoren tot de quarantaine organismen.
Insectenparasitaire rondwormen, de zogenaamde entomopathogene rondwormen, kunnen schadelijke insecten bestrijden, zoals de inzet van Steinernema feltiae bij de bestrijding van de eikenprocessierups,[14] het gebruik van Phasmarhabditis hermaphrodita tegen naaktslakken, bij mais de rondworm Steinernema carpocapsae bij de bestrijding van Helicoverpa zea[88] en de bestrijding van de gegroefde lapsnuitkever met Heterorhabditis bacteriophora. De bacteriën die in een speciale structuur van het aaltje voorkomen doden uiteindelijk de larven en insecten.[89]
Plantenparasitaire aaltjes omvatten verschillende groepen, die ernstige gewasverliezen veroorzaken. De meest voorkomende geslachten met plantenparasitaire aaltjessoorten zijn Aphelenchoides (bladaaltjes), Ditylenchus (stengelaaltje), Globodera (aardappelcystenaaltjes), Heterodera (wit bietencystenaaltje), Longidorus, Meloidogyne (wortelknobbelaaltjes), Nacobbus, Pratylenchus (wortellesieaaltjes), Trichodorus en Xiphinema. Verschillende plantenparasitaire aaltjes veroorzaken schade aan de wortelweefsels, inclusief de vorming van zichtbare gallen, zoals de wortelknobbelaaltjes. Vele plantenparasitaire aaltjes kunnen als er geen plantengastheer aanwezig is, zich ook op schimmelweefsel voortplanten, zoals Aphelenchoides-soorten en de dennenhoutnematode. Laboratoriumtests hebben aangetoond dat Botrytis cinerea en veel Rhizoctonia-soorten gunstige schimmels zijn voor de groei en voortplanting van het chrysantenbladaaltje. Deze schimmels worden ook gebruikt om andere Aphelenchoides-soorten te kweken en te vermeerderen.[90]
Rondwormen kunnen schadelijke virussen overdragen, zoals Xiphinema spp, Longidorus spp, Trichodorus spp. Xiphinema diversicaudatum kan het arabis-mozaïekvirus over brengen bij Fraxinus en het strawberry latent ringspot virus bij aardbeien. Xiphinema americanum is een overbrenger van het tabaksringvlekvirus (tobacco ringspot virus).[91] Xiphinema index draagt het grapevine fanleaf virus over.
Andere rondwormen tasten bomen aan. De belangrijkste vertegenwoordiger van deze groep is de dennenhoutnematode Bursaphelenchus xylophilus, de rondworm dat in dennenbossen voorkomt in Azië en Amerika en in 1999 ontdekt is in Portugal.[92] Deze rondworm maakt gebruik van boktorren van het geslacht Monochamus voor de verspreiding. Om van boom naar boom te komen gebruikt Bursaphelenchus xylophilus de boktor als transportmiddel. Als de kevers in juni tot juli uitkomen kunnen de rondwormen van aangetaste bomen onder de dekschilden en in de ademopeningen meeliften naar gezonde bomen, waar de kever de jonge twijgen en naalden door rijpingsvraat aantast. De rondwormen verlaten op dat moment de boktor en dringen door bastwonden de harskanalen binnen. In het hout vermeerdert de rondworm zich en zorgt voor verstopping van de houtvaten, waardoor de boom verwelkt en dood gaat.[93][94] Het is in Europa een quarantaine organisme.
Parasitaire plantenaaltjes kunnen bestreden worden door nematiciden, vruchtwisseling[95] of gebruik te maken van resistente rassen. Bij het gebruik van resistente rassen kunnen echter pathotypen gaan optreden, zoals bij het aardappelcystenaaltje, waarvan thans vijf pathotypen bestaan. Het blijft dan een race tussen de plantenveredelaar en de aanpassingssnelheid van het aaltje. Ook kunnen plantensoorten als Tagetes patula of andere Tagetes-soorten ingezet worden tegen de wortellesieaaltjes.[96] Het wortelexudaat van de Tagetesplanten thiophene a-terthienyl doodt de aaltjes.[97] Een nadeel hiervan is dat er in het betreffende seizoen geen gewas geteeld kan worden.


Een andere mogelijkheid is het gebruik van natuurlijke antagonisten zoals nematofage schimmels als Clonostachys rosea, waarvan de conidia zich hechten aan de cuticula van het aaltje, ontkiemen en met hun kiemhyfen het aaltje binnendringen en doden.[98] De sikkelvormige, iets schroefvormige mitosporen van de schimmel Harposporium anguillulae worden opgenomen door de mond van de Rhabditis-rondworm. Ze schroeven zich vervolgens met de scherpe punt vast in het farynxweefsel van de rondworm, waar ze kiemen en verder naar binnen groeien en een dikwandig mycelium vormen. Het mycelium lost het weefsel van het verteringskanaal op. Dunwandige hyfen groeien vervolgens door het cuticulum naar buiten en vormen de conidoforen. In de rondworm worden vaak chlamydosporen gevormd. Mortierella globalpina leeft onder andere van het maïswortelknobbelaaltje door de hyfen vast te hechten aan de cuticula, deze te doorboren en vervolgens de cellulaire inhoud van het aaltje te verteren.[99] Engyodontium aranearum en Purpureocillium lilacinum infecteren de eieren van wortelknobbelaaltjes en verteren ze.[100] Ook schimmelsoorten van het geslacht Metacordyceps, zoals Metacordyceps chlamydosporia kunnen parasitaire rondwormeneieren aantasten.[101] Er zijn ook nematofage mijten, zoals Alycus roseus, waarmee onderzoek ter bestrijding van plantenparasitaire aaltjes is gedaan.[102][103]
Chitosan, een natuurlijk biocide, lokt plantenafweerreacties uit waarbij parasitaire cystenaaltjes op de wortels van soja, maïs, suikerbiet, aardappel en tomaat gedood worden, maar niet de niet-parasitaire aaltjes in de bodem.[104] Stomen van de grond, waarbij de grondtemperatuur ten minste moet oplopen tot 80 °C, is een effectieve methode om aaltjes te doden voordat een gewas wordt geplant of gezaaid, maar doodt zowel de schadelijke als de nuttige bodemfauna. CSIRO heeft een 13- tot 14-voudige afname van de populatiedichtheden van Meloidogyne javanica-aaltjes gevonden in percelen met het sareptamosterdras Nemfix (Brassica juncea var. juncea) als groenbemester, dat dient als vanggewas voor de aaltjes.[105] Door de wortels van het vanggewas worden de larven uit de cysten gelokt, maar ze kunnen zich hierop niet vermeerderen. Ook kan bladrammenas als groenbemester ingezet worden tegen het bietencystenaaltje en aardappelmoeheid, maar bladrammenas is een waardplant van het witte (Heterodera schachtii) en het gele bietencystenaaltje (Heterodera trifolii). Daarom moeten er tegen deze aaltjes resistente rassen van bladrammenas worden gebruikt.
Schimmelparasitaire rondwormen, zogenaamde fungivore rondwormen, voeden zich met schimmels, zoals Aphelenchoides-soorten.
Grondmonsteronderzoek
De grond kan onderzocht worden op het voorkomen van plantenparasitaire aaltjes en de te verwachte schade aan het gewas.[106][107][108] Een grondmonster kan in Nederland onderzocht worden door onder andere De Groene Vlieg,[109] Eurofins Agro,[110] HLB (Hilbrands Laboratorium),[111] Nederlandse Algemene Keuringsdienst,[112] Nemacontrol[113] en het Roba Laboratorium.[114]
Bij het grondonderzoek naar vrijlevende aaltjes wordt het grondmonster gespoeld en kruipen de vrijlevende aaltjes door een filter naar een opvangplaats. Vervolgens kunnen ze onder een microscoop op naam gebracht worden en kan hun aantal bepaald worden. Ook kunnen de aaltjes op grond van hun DNA op naam gebracht worden.
Bij het grondonderzoek naar cystenaaltjes wordt het grondmonster met speciale apparatuur gespoeld en worden de resten van het uitgespoelde monster onder een microscoop onderzocht. Hierbij worden het aantal levenskrachtige cysten, het aantal eieren en het aantal larven bepaald.[112]
Bij het grondonderzoek naar wortelknobbelaaltjes worden de grondmonsters geanalyseerd met de incubatietechniek, zodat ook de aaltjes in het plantenweefsel worden meegenomen.
Vijanden
Vijanden van rondwormen zijn protozoa (Apodera vas en Difflugia sp.), amoeben, nematofage schimmels, beerdiertjes,[115] nematovore insecten zoals mijten behorend tot de familie Tydeidae, Ascidae (Protogamasellus mica) en Caloglyphus sp.[103][116][117][118][119] en nematovore rondwormen. Zo at het beertje Macrobiotus richtersi onder laboratorium omstandigheden 61 rondwormen per dag.[120]Apodera vas eet vooral Plectus- en Dorylaimus-rondwormen.[121] Een voorbeeld van een nematovore rondworm is Pristionchus pacificus.
Fotogalerij
 Meloidogyne incognita-aaltje dat een tomatenwortel binnendringt, vergroting 500X
Meloidogyne incognita-aaltje dat een tomatenwortel binnendringt, vergroting 500X Volwassen Caenorhabditis elegans
Volwassen Caenorhabditis elegans Rode lebmaagworm met eieren
Rode lebmaagworm met eieren Model van mannelijke en vrouwelijke rondworm van Trichinella spiralis
Model van mannelijke en vrouwelijke rondworm van Trichinella spiralis Een vrouwelijke Nippostrongylus brasiliensis-aaltje
Een vrouwelijke Nippostrongylus brasiliensis-aaltje Mannelijke Thelazia callipaeda
Mannelijke Thelazia callipaeda Pristionchus pacificus, rondworm met twee verschillende monden
Pristionchus pacificus, rondworm met twee verschillende monden Vrouwtje van Meloidogyne arenaria in een pindawortel
Vrouwtje van Meloidogyne arenaria in een pindawortel Rondwormparasieteneitjes en een Strongyloides stercoralis aaltje
Rondwormparasieteneitjes en een Strongyloides stercoralis aaltje
 Meloidogyne arenaria L2-aalyjes in wortelpunt van de pinda
Meloidogyne arenaria L2-aalyjes in wortelpunt van de pinda L3-aaltje van Phasmarhabditis hermaphrodita
L3-aaltje van Phasmarhabditis hermaphrodita Striae-patroon rond de anus (perinaalpatroon) van een mannelijk Meloidogyne incognita-aaltje
Striae-patroon rond de anus (perinaalpatroon) van een mannelijk Meloidogyne incognita-aaltje Johnstonmawsonia sp.-vrouwtje. A, B: Mond. pijlen geven de onderlippen aan. C: Mond. pijlen geven de tanden aan. D: Excretie porie. a=amfide; b=papillen; c=sublabium (onderkaak).
Johnstonmawsonia sp.-vrouwtje. A, B: Mond. pijlen geven de onderlippen aan. C: Mond. pijlen geven de tanden aan. D: Excretie porie. a=amfide; b=papillen; c=sublabium (onderkaak). Cucullanus bulbosus. A en B: a=amfide, b=papil. A, B: Kop, ventraal en lateraal, C, D: Peribuccale tand mannetje en vrouwtje, E: Staart mannetje, lateraal, F: Staart mannetje, lateraal, G: Staart mannetje lateraal.
Cucullanus bulbosus. A en B: a=amfide, b=papil. A, B: Kop, ventraal en lateraal, C, D: Peribuccale tand mannetje en vrouwtje, E: Staart mannetje, lateraal, F: Staart mannetje, lateraal, G: Staart mannetje lateraal. Mermis nigrescens Fig. 1. Dwarsdoorsnede ongeveer in het midden van het lichaam. Fig. 2. Kopeind. Fig. 3. Staarteind Fig. 4. Enkele spiervezels. Fig. 5. Hersenen (Kopgangliën en zenuwring) Fig: 6. Zenuwen verbonden met de spieren. Fig. 7. Verteringsstelsel. Fig. 8. Uitscheidingsorgaan.
Mermis nigrescens Fig. 1. Dwarsdoorsnede ongeveer in het midden van het lichaam. Fig. 2. Kopeind. Fig. 3. Staarteind Fig. 4. Enkele spiervezels. Fig. 5. Hersenen (Kopgangliën en zenuwring) Fig: 6. Zenuwen verbonden met de spieren. Fig. 7. Verteringsstelsel. Fig. 8. Uitscheidingsorgaan. Kop van Philometra fasciati met papillen
Kop van Philometra fasciati met papillen Philometra protonibeae
Philometra protonibeae Philometra protonibeae, links vrouwtje, rechts mannetje met spiculum
Philometra protonibeae, links vrouwtje, rechts mannetje met spiculum- Torrestrongylus tetradorsalis. Rasterelektronenmicroscopische afbeeldingen.



_-Weisker-.jpg)

.png)
.jpg)


.gif)
.jpg)

.jpg)
.png)
.png)
.jpg)

.png)
.png)
{kind=link}
{kind=link}
{kind=link}
- Onderschrift afbeelding van Torrestrongylus tetradorsalis rasterelektronenmicroscopische afbeeldingen: 13: Subventraal aanzicht van het voorste uiteinde, met laterale afbeeldingen van cefalisch (kop) blaasje; het achterste deel van het cefalisch blaasje lijkt "leeggelopen". 14: Frontaalaanzicht met mondopening, papillen en amfiden. 15: Detail van de vulva met opvallende flap, zijaanzicht. 16: Ventrale weergave van vulva met de basis van de vulvaire flap. 17: Deiride (een sensorische papil als tastorgaan in het laterale cervicale gebied). 18: Zijaanzicht van bursa (staartzakje), afbeelding met bursale stralen. 19: Ventrale weergave van de staart van het vrouwtje, met anus en terminale stekels. Striae zijn zichtbaar. 20: Subterminaal beeld van de staart van het vrouwtje, met alle drie de cuticulaire afbeeldingen en terminaal filament (draadje). Striae zijn zichtbaar.
Zie ook
Externe links
Wikiwand - on
Seamless Wikipedia browsing. On steroids.