Fettsäuresynthese
Art von Synthese Aus Wikipedia, der freien Enzyklopädie
Die biologische Fettsäuresynthese ist ein anaboler, assimilierender Stoffwechselprozess, bei dem Fettsäuren (z. B. zum Zweck der Speicherung von Energie) hergestellt werden. Sie verläuft durch sukzessiven Anbau von Malonyl-CoA an eine initial vorhandene Acetylgruppe, die an Coenzym A gebunden ist. Es lassen sich zwei de novo Fettsäuresynthesen unterscheiden: die cytosolische Fettsäuresynthese (FAS/FAS I) und die mitochondriale Fettsäuresynthese (mtFAS/mtFAS II).
| Übergeordnet |
|---|
| Lipidbiosynthese Metabolismus der Fettsäuren |
| Gene Ontology |
| QuickGO |
Allgemeiner Reaktionsablauf
Zusammenfassung
Kontext
Für die genaue Biosynthese inklusive Strukturformeln siehe Abschnitt Weblinks
Bei Säugetieren und Pilzen bleibt die im Aufbau befindliche Fettsäure bis zur endgültigen Fertigstellung an einem multifunktionellen Enzym, der sogenannten Fettsäure-Synthase Typ I (FAS I),[1] gebunden, die alle sieben Enzymfunktionen vereint. Sie besitzt eine periphere (distale) SH-Gruppe und eine zentrale (proximale) SH-Gruppe an einer Untereinheit des Komplexes, der Acyl-Carrier-Protein-Domäne (ACP). Die einzelnen Enzymfunktionen und ACP sind bei Pflanzen und Bakterien auf verschiedene Proteine verteilt, die sich zu einem Proteinkomplex zusammenlagern. Ebenso existiert bei allen Eukaryoten ein zweiter Fettsäuresyntheseweg in den Mitochondrien, bei welchem die Enzymaktivitäten auf Einzelproteine verteilt sind.[2]
Der Reaktionsweg der cytosolischen Fettsäuresynthese läuft wie folgt ab:
- 1) Acetyl-CoA entsteht in mehreren Prozessen, hauptsächlich durch oxidative Decarboxylierung von Pyruvat in der Glykolyse, durch Abbau von Aminosäuren oder durch β-Oxidation von Fettsäuren, dem umgekehrten zum hier dargestellten Prozess.
- 2)
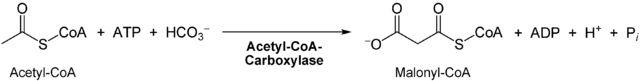
- Carboxylierung (Anlagerung von CO2) von freiem Acetyl-CoA zu Malonyl-CoA außerhalb des Komplexes durch die Acetyl-CoA-Carboxylase (mit der prosthetischen Gruppe Biotin).

- 3)
- Bindung des Malonyl-CoA an die Acyl-Carrier-Protein-Domäne durch die Malonyl-Transferase[3], dabei wird Coenzym A wieder abgespalten.
![{\displaystyle \mathrm {Malonyl{\text{-}}CoA+ACP{\text{-}}SH{\xrightarrow[{ACP-S-Malonyltransferase}]{}}CoA{\text{-}}SH+Malonyl{\text{-}}ACP} }](http://wikimedia.org/api/rest_v1/media/math/render/svg/015a3db00acbd781c6bcb19abb8dddb0abffbbad)
- 4)

- Der Acetylrest kondensiert unter Abspaltung von CO2 mithilfe der Ketoacyl-Synthase[4] an den Malonylrest, es bildet sich eine Acetoacetylgruppe (C4) an der proximalen SH-Gruppe; mit jeder der acht „Zyklen“, die die Reaktion durchläuft, bekommt dieses Zwischenprodukt mehr C-Atome (hat folglich auch einen anderen Namen). In Pflanzen und Bakterien kann auch ein anderes Enzym die Kette verlängern, das Acetyl-CoA direkt als Substrat benutzt (KAS III, EC 2.3.1.180).

- 5)

- Reduktion der Keto-Gruppe des Ketoacylrests an C3 durch die Ketoacyl-ACP-Reduktase[5], das Produkt ist ein Hydroxyacylrest.

- 6)

- Dehydratisierung des Hydroxyacylrests durch die Hydroxyacyl-ACP-Dehydratase[6] zwischen C2 und C3.

- 7)

- Zweite Reduktion der Doppelbindung durch die Enoyl-ACP-Reduktase[7].

- 8) Anlagerung von Malonyl-CoA an die ACP-Domäne (=Schritt 3), anschließend sechsmalige Wiederholung des Zyklus und damit Verlängerung des Acylrests an der proximalen SH-Gruppe.
- 9) Palmitinsäure: C16 wird durch eine Acyl-Hydrolase freigesetzt und dissoziiert sofort zu Palmitat.
Dieser Vorgang wiederholt sich bis zur Fertigstellung, wobei meistens Palmitinsäure durch eine Thiolase hydrolytisch abgespalten wird. Hierbei ist zu beachten, dass die Fettsäure zunächst frei vorliegt. Das Schlüsselenzym der Fettsäuresynthese ist die Acetyl-CoA-Carboxylase, die sowohl allosterisch als auch hormonell reguliert wird.
Die Kettenverlängerung der Fettsäuren wird in der Pflanze durch Elongasen katalysiert.
Dieser Stoffwechselweg spielt beim Menschen unter gewöhnlichen Nahrungsbedingungen eine geringe Rolle, da durch die Nahrung bereits genügend Fette aufgenommen werden. Infolgedessen besteht nicht die Notwendigkeit, aus Kohlenhydraten Fettsäuren aufzubauen. Bei Tieren hat die Fettsäuresynthese noch eine größere Rolle, da diese z. B. eine erhebliche Fettreserve für den Winter bilden müssen.
Ungeradzahlige Fettsäuren
Fettsäureketten mit ungerader Anzahl von Kohlenstoffatomen entstehen, wenn Propionyl-CoA anstelle von Acetyl-CoA als Startmolekül dient. Die Kettenverlängerung findet dann, wie schon bei den geradzahligen Fettsäuren beschrieben, durch sukzessive Reaktionen mit Malonyl-CoA statt.[8]
Verzweigtkettige Fettsäuren
Zusammenfassung
Kontext


Verzweigungen am Alkyl-Ende der Fettsäure ergeben sich, wenn die Synthese mit einem verzweigten Molekül beginnt. Dabei spricht man beim Vorhandensein einer zusätzlichen Methylgruppe am vorletzten Kohlenstoffatom der Hauptkette von einer iso-Fettsäure und am vorvorletzten von einer anteiso-Fettsäure. Iso-Fettsäuren entstehen, wenn die Synthese mit Isovaleryl-CoA oder Isobutyryl-CoA beginnt. Diese Verbindungen leiten sich von den verzweigtkettigen Aminosäuren Leucin beziehungsweise Valin ab. Dagegen entsteht durch den Synthesestart mit 2-Methylbutyryl-CoA, das sich von Isoleucin ableitet, eine anteiso-Fettsäure. Weiter innerhalb der Fettsäurekette können aber ebenfalls Methyl-Verzweigungen während der Synthese erzeugt werden. Das geschieht, wenn die Kettenverlängerung nicht mit Malonyl-CoA, sondern mit Methylmalonyl-CoA durchgeführt wird.[8]
Für etliche Bakterienarten wie die Mykobakterien ist die Synthese von verzweigtkettigen Fettsäuren von großer Bedeutung, da Kettenverzweigungen eine Möglichkeit zur Regulierung der Membranfluidität darstellen. Die Biosynthese der längsten natürlichen Fettsäuren, der Mykolsäuren, ist so umfangreich, dass sie hier nicht detailliert dargestellt werden kann.[9]
Mitochondriale Fettsäuresynthese
Zusammenfassung
Kontext
Neben der Fettsäuresynthese im Cytosol (FAS/FAS I) gibt es bei Eukaryonten auch eine weitere de novo Fettsäuresynthese in den Mitochondrien (mtFAS/mtFASII). Dieser Syntheseweg wurde erstmals 1990 in Neurospora crassa beschrieben.[10][11] Die mitochondriale Fettsäuresynthese ist essentiell für die Zellatmung und die mitochondriale Biogenese.[12] Sie ist auch für das respiratorische Wachstum in Hefen und für das embryonale Überleben in Säugetieren erforderlich.[13]
Reaktionsablauf
_pathway.svg)
Der mtFAS-Stoffwechselweg besteht aus mindestens sechs einzeln vorhandenen Enzymen, die alle von separaten Genen kodiert werden.[14] Dies unterscheidet ihn von der cytosolischen Fettsäuresynthese, bei der das multifunktionelle Enzym Fettsäuresynthase (FASN) alle enzymatischen Aktivitäten in einer einzigen Polypeptidkette enthält und von einem einzigen Gen kodiert wird.[14] Trotz dieses strukturellen Unterschieds verwenden mtFAS und FAS die gleiche Chemie, um Fettsäuren zu bilden.[10]
Bei der mtFAS dient das mitochondriale Acyl-Carrier-Protein (ACP) als lösliches Gerüstprotein in der mitochondrialen Matrix, das die wachsenden Fett-Acyl-Ketten kovalent an sich bindet.[10] Malonyl-CoA, das von mtACC1 (einer mitochondrialen Isoform der Acetyl-CoA-Carboxylase 1) aus Acetyl-CoA und von Acyl-CoA-Synthetase-Familienmitglied 3 (ACSF3) aus Malonsäure gebildet wird, dient als Kettenverlängerungseinheit.[15][16] Die genaue mitochondriale Quelle für Malonyl-CoA ist jedoch noch nicht abschließend geklärt.[17]
In jeder Runde der Kettenverlängerung wird zunächst Malonyl-CoA durch Malonyl-CoA:ACP-Transacylase (MCAT) auf ACP übertragen, um Malonyl-ACP zu bilden, das dann mit dem wachsenden Acyl-ACP (mit Acetyl-ACP in der ersten Runde) kondensiert, katalysiert durch 3-Oxoacyl-ACP-Synthase (OXSM), wobei CO2 freigesetzt und die Kette um zwei Kohlenstoffe verlängert wird.[14] Anschließend durchläuft die neu verlängerte Fettacylkette an ACP (3-Ketoacyl-ACP) eine Reduktion durch Estradiol-17β-Dehydrogenase 8 (HSD17B8) und Carbonylreduktase 4 (CBR4), eine Dehydratisierung durch 3-Hydroxyacyl-ACP-Dehydratase 2 (HTD2), und eine weitere Reduktion durch trans-2-Enoyl-CoA-Reduktase (MECR), wodurch wieder eine gesättigte Fettsäure an ACP (Acyl-ACP) entsteht, die dann als Substrat für die nächste Verlängerungsrunde zur Verfügung steht.[14]
Diese Schritte wiederholen sich, bis eine achtkettige, gesättigte Fettsäure an ACP – bekannt als Octanoyl-ACP (C8) – gebildet ist.[10] An diesem Punkt kann diese an ACP gebundene mittelkettige Fettsäure entweder den mtFAS-Stoffwechselpfad verlassen oder für die weitere Verlängerung zu langkettigen Fettsäuren (C14-C16) auf diesem verbleiben.[10] Da bei keiner Tierart eine mitochondriale Thioesterase identifiziert wurde, bleibt das Endprodukt der mtFAS an ACP gebunden und wird nicht als freie Fettsäure freigesetzt.[10]
Funktion
Die mitochondriale Fettsäuresynthese spielt eine entscheidende Rolle im zellulären Energiestoffwechsel, indem sie Octanoyl-ACP (C8) erzeugt, das als direkter Vorläufer für die Biosynthese von Liponsäure dient.[18] Liponsäure ist ein essentieller Cofaktor, der in einem als Lipoylierung bezeichneten Prozess kovalent an spezifische Lysinreste von Zielenzymen gebunden wird.[19] Diese posttranslationale Modifikation ist wesentlich für die Aktivität wichtiger mitochondrialer Enzymkomplexe, namentlich des Pyruvat-Dehydrogenase-Komplexes (PDC), des α-Ketoglutarat-Dehydrogenase-Komplexes (OGDC), des 2-Oxoadipat-Dehydrogenase-Komplexes (2-OADHC), des verzweigtkettigen α-Ketosäure-Dehydrogenase-Komplexes (BCKDC) und des Glycine-Cleavage-System (GCS).[19][20]
Parallel dazu stellen mtFAS und seine Acyl-ACP-Produkte einen metabolischen Rückkopplungsmechanismus dar, der den Verbrauch von Acetyl-CoA in den Mitochondrien reguliert und so die Lipidsynthese mit der allgemeinen Stoffwechselkontrolle verbindet.[17]
Neben Octanoyl-ACP produziert mtFAS auch längerkettige Acyl-ACP-Spezies wie Myristoyl-ACP (C14) und Palmitoyl-ACP (C16), die mit Mitgliedern der Leucin-Tyrosin-Arginin-Motiv-Proteinfamilie (LYRM) interagieren.[10] Diese LYRM-Proteine sind für die korrekte Assemblierung und die Stabilität der Elektronentransportketten-Komplexe (ETC) und für die Biogenese von Eisen-Schwefel-Clustern (Fe-S) in den Mitochondrien entscheidend.[10]
Zusätzlich zu diesen enzymatischen und strukturellen Aufgaben wird die mtFAS auch als Mediator der intrazellulären Signaltransduktion in Verbindung gebracht.[21] Dies wird durch die Beobachtung gestützt, dass der Gehalt an bioaktiven Lipiden – wie Lysophospholipiden und Sphingolipiden – mit der mtFAS-Aktivität korreliert.[21] So führt beispielsweise der Knock-down von ACP zu einer Verringerung des Ceramidgehaltes, während der Verlust des terminalen mtFAS-Enzyms MECR zu einer Ceramidanhäufung führt.[21][22]
Eine wichtige Funktion von mtFAS ist die Regulierung des Immunzellstoffwechsels. CRISPR/Cas9-Screens haben die mtFAS-Gene – insbesondere Mecr, Mcat und Oxsm – als Schlüsselregulatoren des T-Zell-Stoffwechsels identifiziert.[23] Während MECR für die Aufrechterhaltung der naiven T-Zellen nicht erforderlich ist, beeinträchtigt sein Verlust in aktivierten T-Zellen die Proliferation, das Überleben und die Differenzierung.[23] MECR-Mangel stört die mitochondriale Atmung, verändert die Citratzyklus-Aktivität und erhöht die Ferroptose-Empfindlichkeit, was letztlich die T-Zell-Fitness und die Entzündungskapazität reduziert.[23]
Krankheiten
Störungen im mtFASII-Pfad führen zu den folgenden Stoffwechselerkrankungen:
- ACSF3: Kombinierte Malon- und Methylmalonazidurie (CMAMMA)[24]
- MECR: mitochondriale Enoyl-CoA-Reduktase-Protein-assoziierte Neurodegeneration (MEPAN)[24]
Vergleich der cytosolischen und mitochondrialen Fettsäuresynthese
Zusammenfassung
Kontext
Nachfolgend werden Gemeinsamkeiten und Unterschiede zwischen der cytosolischen und mitochondrialen Fettsäuresynthese aufgezeigt:
| Merkmal | Cytosolische Fettsäuresynthese (FAS/FAS I) | Mitochondriale Fettsäuresynthese (mtFAS/mtFAS II) | |
|---|---|---|---|
| Ort der Synthese | Cytosol | Mitochondriale Matrix[10] | |
| Enzymsystem | FAS Typ I (multifunktionelles Enzym) | FAS Typ II (einzelne Enzyme)[10] | |
| Regulation | Schlüsselenzym | Acetyl-CoA-Carboxylase | Unbekannt |
| Aktivierung | Allosterisch: Citrat | Unbekannt | |
| Hemmung | Allosterisch: Palmitoyl-CoA | Unbekannt | |
| Primer | Acetyl-CoA (aus Mitochondrien über Citrat-Malat-Shuttle) | Acetyl-CoA (direkt in der Mitochondrienmatrix vorhanden), Malonsäure[10] | |
| Verlängerungseinheiten | Malonyl-CoA (aus der Carboxylierung von Acetyl-CoA) | Malonyl-CoA (hauptsächlich aus der Carboxylierung von Acetyl-CoA, aber auch aus der Thioesterifizierung von Malonsäure) | |
| Cofaktoren | Reduktionsmittel | NADPH | NADPH |
| Andere | ATP, Biotin (beide für die Umwandlung zu Malonyl-CoA) | ATP, Biotin (beide auch für Malonyl-CoA) | |
| Thioesterase | Vorhanden im Cytosol | Keine bekannt in den Mitochondrien[10] | |
| Endprodukt(e) | Hauptsächlich Palmitat (C16:0) | Octanoyl-ACP (C8), Myristoyl-ACP (C14), Palmitoyl-ACP (C16)[10] | |
| Funktion | Lipidspeicherung, Energiegleichgewicht, Membranstruktur | Vorstufen für Cofaktoren, wie Liponsäure (für PDH-Komplex, αKGDH-Komplex, 2-Oxoadipat-Dehydrogenase-Komplex, BCKDH-Komplex und Glycine-Cleavage-System);[18][10] Assemblierung der Elektronentransportkette (ETC);[10] Biogenese von Eisen-Schwefel-Clustern;[10] Rolle im Ceramid-Stoffwechsel[10] | |
| Teilnahme an Lipidsynthese | Zentrale Rolle bei der de novo Lipogenese | Nur ergänzende Rolle | |
| Phylogenetische Ähnlichkeiten | Eukaryoten-spezifisch | Bakterienähnlich (evolutionär konserviert) | |
Fettsäuresynthese bei Pflanzen
Bei Pflanzen läuft die Fettsäuresynthese nur in den Plastiden ab, bei grünen Pflanzenzellen in den Chloroplasten, sonst in den Chromoplasten, Leukoplasten oder Proplastiden. Es laufen dieselben Reaktionen wie im Cytosol ab, jedoch werden nur Fettsäuren bis C18 synthetisiert. Diese können maximal eine Doppelbindung durch eine lösliche Desaturase des Stromas erhalten. Nach Transport ins glatte endoplasmatische Retikulum (sER) erfolgt die Kettenverlängerung, im sER können durch membrangebundene Desaturasen weitere Doppelbindungen eingebaut werden. Der Abbau der Fettsäuren erfolgt bei Pflanzen nicht in den Mitochondrien, sondern nur in Peroxisomen.
Langkettige Fettsäuren sind in Pflanzen der Ausgangspunkt für die Biosynthese von Wachs und Cutin, welche beide für den Schutz vor der Umgebung unentbehrlich sind.
Siehe auch
Literatur
- Florian Horn: Biochemie des Menschen. Das Lehrbuch für das Medizinstudium. 3. Auflage. Thieme, 2005, ISBN 3-13-130883-4.
- Jeremy M. Berg, John L. Tymoczko, Lubert Stryer: Biochemie. 5. Auflage. Spektrum Akademischer Verlag, Heidelberg 2003, ISBN 3-8274-1303-6.
- Peter Sitte, Elmar Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger. 35. Auflage. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X.
Weblinks
Wikibooks: Biosynthese gesättigter Fettsäuren – Lern- und Lehrmaterialien
- Animation des Reaktionsablaufs
- Gopinathrao/reactome.org: Fatty Acyl-CoA Biosynthesis
Einzelnachweise
Wikiwand - on
Seamless Wikipedia browsing. On steroids.