チミジンキナーゼ
ウィキペディアから

チミジンキナーゼ(英: thymidine kinase、略称: TK、EC 2.7.1.21)は酵素(ホスホトランスフェラーゼ、キナーゼ)であり、より具体的には2'-デオキシチミジンキナーゼ(2'-deoxythymidine kinase)、ATP-チミジン 5'-ホスホトランスフェラーゼ(ATP-thymidine 5'-phosphotransferase)である[2][3]。大部分の細胞に存在し、哺乳類細胞にはTK1、TK2という2種類が存在する。特定のウイルスもチミジンキナーゼを発現するための遺伝情報を保有している。チミジンキナーゼは次の反応を触媒する。
| Thymidine kinase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 チミジンと複合体を形成したUreaplasma urealyticum由来チミジンキナーゼ四量体の結晶構造(各単量体はシアン、緑、赤、マゼンタで示されている)。チミジンは空間充填モデルで示されている[1]。 | |||||||||
| 識別子 | |||||||||
| EC番号 | 2.7.1.21 | ||||||||
| CAS登録番号 | 9002-06-6 | ||||||||
| データベース | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB構造 | RCSB PDB PDBj PDBe PDBsum | ||||||||
| 遺伝子オントロジー | AmiGO / QuickGO | ||||||||
| |||||||||
チミジンキナーゼはチミジンをDNAに導入するための反応の一部を構成しており、DNA合成、そして細胞分裂に重要である。また、チミジンキナーゼは多くの抗ウイルス薬の作用に必要である。モノクローナル抗体産生のためのハイブリドーマ細胞株の選別にも用いられる。臨床化学においては、悪性腫瘍、主に血液の腫瘍の診断、治療の制御、治療後フォローアップにおける増殖のマーカーとして利用される。
歴史
DNAへのチミジンの取り込みは1950年ごろに示された[4]。その後、この取り込みに先立ってリン酸化が行われることが示され[5]、この反応を担う酵素の精製と特性解析が1960年ごろに行われた[6][7]
アイソザイム
要約
視点
哺乳類にはTK1、TK2と呼ばれる2つのアイソザイムが存在する。TK1はまず胚組織で見つかり、一方TK2は成体組織に豊富に存在することが発見されたため、当初はそれぞれfetal(胎児型)TK、adult(成体型)TKと呼ばれていた。その後、TK1は細胞分裂前にのみ(細胞周期依存的に)細胞質に存在し[8][9]、一方TK2は細胞周期非依存的にミトコンドリアに位置していることが示された[10][11]。2つのアイソザイムは反応速度論的に異なり、異なる阻害剤によって阻害される。
ウイルスのチミジンキナーゼは哺乳類の酵素とは構造的にも生化学的にも全く異なり、哺乳類の酵素を阻害しない阻害剤によって阻害される[12][13][14]。ヒトの2つのアイソザイムの遺伝子の位置は1970年代半ばに特定され[15][16]、TK1の遺伝子のクローニングと配列決定が行われた[17]。対応するタンパク質の大きさは約25 kDaである。組織中では通常は約50 kDaの二量体として存在し、ATPによって活性化されると約100 kDaの四量体を形成する[18]。一方で、血中に存在する活性型酵素の主要画分は約730 kDaであり、おそらく他のタンパク質と複合体を形成している。この複合体はどの低分子量形態よりも安定性が高く、高い特異的活性を示す[19][20]。
組換え型TK1ではこうした形での活性化と四量体への変化は起こらず、細胞内の酵素は合成後に修飾が行われていることが示されている[18][21][22]。
TK1は細胞周期のS期の間に合成される。細胞分裂が完了するとTK1は細胞内で分解され、細胞分裂が正常に終結した場合には体液へ流出することはない[23][24][25][26]。細胞内でのチミジンキナーゼの作用にはフィードバック調節が存在し、さらなるリン酸化の産物であるチミジン三リン酸(TTP)はチミジンキナーゼの阻害剤として作用する[21]。その結果、核酸合成のために利用可能なTTPの量の均衡が得られ、系が過飽和に達することはない。チミジンの非毒性アナログである5'-アミノチミジンはこの調節機構に干渉し、それによって抗がん剤として用いられるチミジンアナログの細胞毒性を高める[27][28][29][30][31][32][33]。チミジンやチミジンアナログのリン酸化の反応速度論は複雑であり、部分的な理解しか得られていない。チミジンからTTPへの全体的なリン酸化はミカエリス・メンテン型の速度論には従わず、チミジンやウリジンのさまざまなリン酸化物が互いのリン酸化に干渉している[34]。生物種によってTKの速度論は互いに異なり、また特定の種の中でもその形態(単量体、二量体、四量体、血清型)によって速度論的特性が異なる。
ウイルス特異的チミジンキナーゼは単純ヘルペスウイルス、水痘帯状疱疹ウイルス、EBウイルスで同定されている[35][36][37][38][39][40][41]。
 + ATP --->
+ ATP --->
 + ADP
+ ADP
チミジンはATPと反応し、チミジン一リン酸とADPが生じる。
機能
チミジンキナーゼによって触媒される反応の産物であるチミジン一リン酸は、チミジル酸キナーゼによってリン酸化されてチミジン二リン酸となり、ヌクレオシド二リン酸キナーゼによってさらにリン酸化されてチミジン三リン酸となる。チミジン三リン酸は、DNAポリメラーゼと相補的DNA分子(レトロウイルスの逆転写酵素の場合にはRNA分子)によって触媒される反応によってDNA分子に組み込まれる。
チミジン一リン酸は細胞内の他の反応によっても産生され、一例としてチミジンとは無関係な代謝経路の産物であるデオキシウリジン一リン酸のチミジル酸シンターゼによるメチル化によって産生される。DNA修復のためのチミジン一リン酸の供給にはこの経路で十分である。一方で細胞が分裂を行おうとするときには、DNAの新たなセットが必要であり、チミジン一リン酸などのビルディングブロックの需要が増大する。細胞は分裂時に必要な酵素の一部を合成することで分裂に備える。これらは通常は細胞内に存在せず、ダウンレギュレーションや分解が行われている。こうした酵素はサルベージ酵素(salvage enzyme)と呼ばれている。TK1はサルベージ酵素の一例であるが、TK2やチミジル酸シンターゼは細胞周期依存性を示さない[42][43][44][45][46][47][48][49][50][51][52]。
欠乏症
TK2はミトコンドリアDNAの合成のために利用される。TK2をコードする遺伝子の変異はミオパチー型のミトコンドリアDNA枯渇症候群の原因となる。TK2の欠乏は、酸化ストレスによるミトコンドリアTK2のS-グルタチオン化とタンパク質分解を原因とするものである可能性もある[53]。TK1の欠乏を原因とする疾患は知られておらず、おそらくTK1をコードする遺伝子の欠陥は胎生致死となるためである。
チミジンキナーゼの進化
TK1合成後の修飾による四量体の形成は酵素活性を高め、酵素活性の調節機構となっていることが示唆されている。四量体の形成はタマホコリカビ属系統の分岐後の動物系統で観察される。そのDNA合成の微調節への利用は、温血脊椎動物が脊椎動物から分岐した後に確立されたことが示唆されている[54]。また、後生動物におけるTK様酵素ファミリーの進化の研究も行われている[55]。
種分布
チミジンキナーゼは、動物[56][57][58][59][60][61][62] 、植物[63][64] 、一部の細菌、古細菌[65][66][67]、ウイルスに存在する。ポックスウイルス[68][69]、アフリカ豚熱ウイルス[70]、単純ヘルペスウイルス[14][35][36][37][38][71][72][73]、水痘・帯状疱疹ウイル[39][74][75]、EBウイルス[40]由来のチミジンキナーゼが同定されており、程度は異なるものの特性解析がなされている。ウイルス由来の酵素は他の生物のものとは特性が異なる[14]。チミジンキナーゼは菌類には存在しない[66][76][77][78]。
応用
要約
視点
分裂細胞の特定
生化学研究においてチミジンキナーゼが初めて間接的に利用されたのは、放射性標識チミジンの組み込み、そしてその後の放射能測定またはオートラジオグラフィーによる分裂細胞の特定である。その際には、増殖培地中にトリチウム化チミジンを添加することで利用された[79]。
活動的な腫瘍に対するPETスキャン
フルオロチミジンはチミジンのアナログである。その取り込みはTK1によって調節されており、急速に増殖している腫瘍組織に選択的に取り込まれる。そのため、ポジトロン断層法(PET)で用いられる陽電子放出核種であるフッ素18(18F)で標識された18F-フルオロチミジンは活発な腫瘍増殖のPETイメージングに有用であり、より広く利用されているフルデオキシグルコース (18F)と比較してより良好な結果が得られる[80][81][82][83][84][85]。臨床試験の比較に有用な標準化プロトコルが提案されている[86]。
ハイブリドーマの選別
ハイブリドーマは、腫瘍細胞(無限に分裂することができる)と免疫グロブリン産生リンパ球(形質細胞)との融合によって得られる細胞である。ハイブリドーマは特定の特異性を持った免疫グロブリン(モノクローナル抗体)の大量生産を行うために増殖させることができる。その過程で問題となることの1つは、細胞融合処理後に大量の非融合細胞の中からハイブリドーマを単離することである。この問題を解決する一般的手法の1つは、チミジンキナーゼ陰性(TK-)腫瘍細胞株を融合に用いることである。TK-細胞は、チミジンキナーゼ陽性(TK+)細胞を死滅させるチミジンアナログの存在下で腫瘍細胞株を生育することで得られる。その後、TK-細胞を増殖し、TK+形質細胞との融合に利用する。融合後、メトトレキサート[87]もしくはアミノプテリン[88]含有培地で生育を行う。これらはジヒドロ葉酸レダクターゼを阻害し、チミジン一リン酸のde novo合成を遮断する。こうした培地で広く用いられているのはHAT培地であり、ヒポキサンチン、アミノプテリン、チミジンを含有する。TK-細胞に由来する非融合細胞はチミジン一リン酸の供給源を失うため、死滅する。また、リンパ球は不死化していないため、こちらも最終的には死滅し、TK-腫瘍細胞由来の不死性と形質細胞由来のチミジンキナーゼを併せ持つハイブリドーマのみが生存する。その後、目的の抗体を産生するハイブリドーマが選択され、モノクローナル抗体の生産のために培養される[89][90][91][92][93]。ハイブリドーマ細胞は、HGPRT遺伝子に対して同様の原理を用いることでも単離することができる。HGPRTはサルベージ経路のGMP合成に必要なIMPを合成する。
染色体構造の研究
DNA繊維の分子コーミング(molecular combing)は、出芽酵母Saccharomyces cerevisiaeの染色体構造をモニターするために利用される。この手法によって個々の分子のDNA複製プロファイルを得ることができるが、この手法を用いるためにはチミジンキナーゼを発現する酵母株が必要である(野生型酵母はチミジンキナーゼを発現しない)。そのため、チミジンキナーゼをコードする遺伝子をゲノムに組み込むことが必要となる[94]。
臨床化学
→詳細は「臨床化学におけるチミジンキナーゼ」を参照
チミジンキナーゼTK1は細胞分裂に備えた細胞のみに存在するサルベージ酵素である。この酵素は正常な分裂を行っている細胞から遊離することはなく、細胞分裂後に不要となったタンパク質を分解する特別な機構が備わっている[8]。そのため、正常な被験者では血清や血漿中のチミジンキナーゼの量は極めて低い。一方で腫瘍細胞からは酵素が血中に放出され、おそらくこの現象は死んだもしくは死にかかっている腫瘍細胞の破壊と関係している。そのため、血清中のチミジンキナーゼ濃度は悪性増殖の尺度となり、また腫瘍の悪性度の間接的な尺度となる。
治療応用
一部の医薬品は分裂中の細胞を特異的な標的としており、こうした医薬品は腫瘍やウイルス性疾患に対して使用される。こうした疾患の細胞は正常細胞よりも極めて高い頻度で複製を行うためであるが、過剰な細胞複製と関係した非悪性疾患(乾癬など)に対しても使用される。チミジンアナログの抗ウイルス・抗がん作用の少なくとも一部は、ミトコンドリア型チミジンキナーゼ(TK2)のダウンレギュレーションによるものであることが示唆されている[95]。
細胞分裂阻害薬
チミジン代謝を標的とした、チミジンキナーゼと関係するさまざまなクラスの薬剤が、がんと関係した細胞分裂の制御に利用されている[96][97][98][99][100][101]。チェーンターミネーター(chain terminator)は伸長中のDNA鎖に組み込まれるチミジンアナログであり、鎖がそれ以上伸長することがないような修飾が施されている。この種の薬剤はチミジンアナログであるため、迅速に5'-一リン酸化が行われる。一リン酸化物はさらに対応する三リン酸化物にまでリン酸化され、伸長中のDNA鎖に組み込まれる。このアナログには、鎖の伸長の継続に必要な3'位のヒドロキシル基が存在しない。ジドブジン(AZT)では3'-ヒドロキシル基はアジド基で置換されており[34][100]、スタブジン(サニルブジン)では置換ではなく除去されている[102][103]。AZTは血清中のチミジンキナーゼ活性の測定法の1つで基質として利用される[104]。AIDSで生じる一般的な症状の1つはリンパ腫であり、チミジンキナーゼ測定の最も重要な診断的用途はリンパ腫のモニタリングであるが、AZTはHIV感染におけるHAART療法の標準的な構成要素であるため、この手法に干渉する可能性がある。
その他のチミジンアナログとして、例えばイドクスウリジンはその後の複製サイクルにおける塩基対形成を遮断し、DNA鎖の欠陥を引き起こす[105]。この薬剤は悪性細胞のアポトーシスを引き起こすため、放射性標識と組み合わせられる可能性もある[106]。
- チミジンキナーゼの基質アナログの化学構造
 ジドブジン
ジドブジン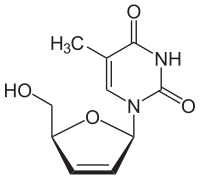 スタブジン
スタブジン イドクスウリジン
イドクスウリジン アシクロビル
アシクロビル ガンシクロビル
ガンシクロビル





抗ウイルス薬
アシクロビルやガンシクロビルなど一部の抗ウイルス薬やその他のヌクレオシドアナログは、ウイルスのチミジンキナーゼの特異性がヒトのチミジンキナーゼとは異なることを利用する[13]。こうした薬剤はプロドラッグとして作用し、それ自体は有毒ではないが、ウイルスのチミジンキナーゼによってリン酸化されることで毒性薬剤へと変換される。そのため、ウイルスが感染した細胞では毒性の高い三リン酸化物が産生され、細胞死が引き起こされる。対照的に、ヒトのチミジンキナーゼの基質特異性はより狭いため、プロドラッグのリン酸化と活性化を行うことはない。このようにして、ウイルスが感染した細胞のみが薬剤に対して感受性となる。こうした薬剤は特異的チミジンキナーゼを持つヘルペスウイルス群に対してのみ有効である[107][108]。このタイプの薬剤で治療を行った患者では、抗ウイルス薬耐性が高頻度で観察される。単純ヘルペスウイルスや水痘帯状疱疹ウイルスのチミジンキナーゼ遺伝子は急速な遺伝的多様性を示し、その配列決定は抗ウイルス薬耐性の診断に有用である[14][75]。
天然痘は1979年12月にWHOによって根絶が宣言され、予防接種プログラムは終結している。そのため、事故もしくは生物兵器としての使用によって疾患が再興した場合に人々は未保護のまま曝されることとなり、制御困難なエピデミックとなる可能性がある。承認されている唯一の天然痘ワクチンであるワクシニアウイルスは重大な副作用が発生する可能性があるため、集団接種による天然痘エピデミックへの対抗には困難が伴う。一部の政府はこうした可能性に備えて天然痘ワクチンの備蓄を行っているが、特異的で有効な抗ウイルス薬の開発が優先される。可能なアプローチの1つとして、ヘルペスウイルスに用いられる薬剤と同様に、ポックスウイルスのチミジンキナーゼの特異性を利用することが考えられる。予測される困難さの1つとして、ポックスウイルスのチミジンキナーゼはヒトのチミジンキナーゼと同じファミリーに属し、そのため化学的特性がより類似していることが挙げられる。抗ウイルス薬候補の探索のためポックスウイルスのチミジンキナーゼの構造決定が行われているが[69]、現在のところポックスウイルスに対して使用可能な抗ウイルス薬は得られていない。
遺伝子治療における自殺遺伝子として
ヘルペスウイルスのチミジンキナーゼ遺伝子は「自殺遺伝子」として、遺伝子治療実験における安全装置として利用されている。この遺伝子を発現する細胞はガンシクロビルを用いて死滅させることができ、このことは組換え遺伝子が無制御な細胞成長をもたらすような変異(挿入変異)を引き起こした場合に都合が良い。こうした改変細胞で産生される細胞傷害性産物が周囲へ拡散することで周辺の細胞も同様にガンシクロビル感受性となる可能性があり、この現象は「バイスタンダー効果」として知られる。このアプローチは動物モデルでのがんの治療に利用されており、遺伝子を発現する悪性細胞がわずか10%程度であっても腫瘍を死滅させる可能性がある点で優位性がある[109][110][111][112][113][114][115][116][117][118][119][120][121][122]。トマトのチミジンキナーゼとAZTを用いた同様の系の試みもなされている[123][124]。さらに、チミジンキナーゼは造血幹細胞移植時の移植片対宿主病に対処する自殺遺伝子としても利用されており、Zalmoxisの名称で2016年にEUで条件付き承認が行われた[125]。
腫瘍マーカー遺伝子
チミジンキナーゼの利用法として、一部の腫瘍細胞に存在し、正常細胞には存在しないもの(腫瘍マーカー)を活用したものがある。腫瘍マーカーの例としては、癌胎児性抗原(CEA)やα-フェトプロテイン(AFP)が挙げられる。こうした腫瘍マーカー遺伝子をチミジンキナーゼのプロモーターとして利用すると、腫瘍マーカーを発現する細胞ではチミジンキナーゼが活性化されるが、正常細胞では活性化されないため、ガンシクロビル治療によって腫瘍細胞のみを死滅させることができる[126][127][128][129][130][131]。こうした遺伝子治療ベースのアプローチは未だ実験段階であり、腫瘍細胞を標的とした遺伝子導入に関する問題は完全に解消されてはいない。
腫瘍に対する中性子捕捉療法
ホウ素含有チミジンアナログの取り込みは脳腫瘍に対する中性子捕捉療法への利用が提案されており、動物モデルでの試行がなされている。これまでに非常に多くのホウ素含有チミジン誘導体が記載されている[132][133][134][135][136][137][138][139][140][141][142][143][144][145][146][147][148]。
抗寄生虫薬
寄生虫のゲノムにチミジンキナーゼを導入することでBrdUの組み込みが可能となり、寄生虫ゲノムの複製の高感度の指標としても利用できるとともに、寄生虫はこの薬剤の毒性に対して感受性となる[149]。
測定
要約
視点
血清や血漿中のTKの測定
→詳細は「臨床化学におけるチミジンキナーゼ」を参照
血清や血漿中のチミジンキナーゼ濃度は、主に酵素活性アッセイを用いて測定される。市販のアッセイでは、血清試料と基質アナログをインキュベーションし、形成された産物の量を測定する[71][72][73][104][150][151][152][153][154][155]。イムノアッセイによるチミジンキナーゼタンパク質の直接的定量も行われる[156][157][158][159][160]。この手法で決定されたチミジンキナーゼの量は、酵素活性から得られる値との良い相関は得られない。その理由の1つはイムノアッセイによって同定される血清型TK1の多くが酵素活性を持たないためである[20][161]。このことは、イムノアッセイの感度がより高くなる可能性がある固形腫瘍の場合にも当てはまる[162][163]。
組織中のTKの測定
→詳細は「臨床化学におけるチミジンキナーゼ」を参照
組織試料中のチミジンキナーゼは組織からの抽出後に測定される。抽出やアッセイの標準的手法は開発されておらず、細胞や組織の抽出物中のTKの定量法は特定の臨床的疑問に関して検証されたものはないが、いくつかの評価が行われている[164][165]。細胞抽出物中のTK2の特異的定量法の1つとして、基質アナログである5-ブロモビニル-2'-デオキシウリジンを用いる手法が開発されている[166]。ここからいくつかの研究について記載するが、用いられた手法や結果の報告法は研究によって大きく異なるため、研究間の比較は不可能である。発生中の胚組織中のTK1レベルは、より後の段階の対応する組織よりも高い[167][168][169]。単球増多症における末梢リンパ球[170]や悪性貧血時の骨髄[171][172]など、特定の非悪性疾患においても細胞や組織内のTK値が劇的に上昇する場合がある。
免疫染色
免疫組織化学的検出のために抗チミジンキナーゼ抗体を利用することができる[173]。チミジンキナーゼに対する染色は、ステージ2乳がんの患者を特定するための信頼性の高い手法である。チミジンキナーゼに対する染色とKi-67に対する染色を組み合わせることで、最も多くの患者を特定することができる[174][175]。この手法は、肺がん[176][177]、大腸がん[178]、腎細胞がん[179]においても有効性が確認されている。
蛍光染色
2'-デオキシ-2',2'-ジフルオロ-5-エチニルウリジン(dF-EdU)は単純ヘルペスウイルスのチミジンキナーゼに結合するが、立体障害のためヒトのチミジンキナーゼには結合しない。この試薬を蛍光アジドと共に用いることで、感染細胞のみに蛍光を引き起こすことができる。そのため、この基質アナログは感染細胞特異的な染色を可能にする[180]。
出典
関連文献
関連項目
外部リンク
Wikiwand - on
Seamless Wikipedia browsing. On steroids.