トップQs
タイムライン
チャット
視点
Gタンパク質共役受容体
ウィキペディアから

Remove ads
Gタンパク質共役受容体(ジータンパクしつきょうやくじゅようたい、英: G protein-coupled receptor、GPCR)は、生体に存在する受容体の形式の1つである。様々な機能を持ったGタンパク質共役受容体が見られ、既知のタンパク質の中では最大のスーパーファミリーを形成している。別名としてGタンパク質結合受容体、あるいは細胞膜を7回貫通する特徴的な構造から

Remove ads
歴史
1986年、網膜に存在する光受容体ロドプシンと、心臓に存在するβ2アドレナリン受容体(β2AR)が発見された。この2つの分子は、限局している領域も機序も異なるタンパク質ではあったが、「膜を7回貫通している」という構造的な共通点が存在した。最初のGタンパク質共役受容体の結晶構造が2000年にウシの網膜から精製したロドプシンで決定され[4]。それまで同じ7回膜貫通タンパク質であるバクテリオロドプシンの光駆動水素イオンポンプ構造モデルより、複雑な折れ曲りをもち、さらに7回貫通ヘリックスの後に短い8番目のヘリックスがあるGタンパク質共役受容体に共通の構造が明らかになった。
そして、2007年にブライアン・コビルカとReymond Stevesたちにより、医薬品標的として初めて、昆虫細胞で発現したβ2-アドレナリン受容体T4リゾチームキメラタンパク質の結晶構造が決定され[5]、2011年の極めて精緻な結晶化技術を駆使して、活性化状態のβ2-アドレナリン受容体とG-タンパク質三量体の結晶構造がコビルカたちのグループによって決定され、コビルカのボスでGタンパク質共役受容体の機能解析の専門家であるロバート・レフコウィッツとともに翌年のノーベル化学賞を受賞した[6]。現在までにロドプシン・ファミリーであるクラスAはもちろんクラスB、クラスC、クラスFの立体構造がすでに解かれている。
2018年に発表された医薬産業政策研究所の調査によると、GPCR標的の新薬開発品目数は、開発薬全体の20%(253/1265)を占め、主要な標的分子群のひとつとなっている[7]。
Remove ads
分類
Gタンパク質共役受容体は、アミノ酸配列や機能の類似に基づいて6つのクラスに分類されている[8][9][10][11]。
- クラス A
- ロドプシン様受容体
- クラス B
- セクレチン受容体ファミリー
- クラス C
- 代謝型グルタミン酸受容体
- クラス D
- 真菌の接合因子受容体
- クラス E
- サイクリックAMP(cAMP)受容体
- クラス F
- Frizzled、Smoothened
ロドプシン様クラスA受容体は、さらに19のサブグループに分けられている(A1-A19)[12]。最近、GRAFS という別の分類法が提案された。これは、代謝型グルタミン酸受容体・ロドプシン・接着因子受容体・フリズルド/苦味受容体・セクレチン受容体の5つに分類するものである[13]。
ヒトゲノムには約800種類のGタンパク質共役受容体がコードされており、これらはホルモンや成長因子をはじめとする内因性リガンドを認識する。ヒトのGタンパク質共役受容体のうち半数は臭い受容体であり、約30種類は、まだリガンドや働きが分かっていないオーファン受容体である。
Remove ads
種類
- ムスカリン性アセチルコリン受容体
- 神経伝達物質アセチルコリンの受容体の1種で、キノコ由来の毒物ムスカリンを結合する特徴がある。
- アデノシン受容体
- 神経伝達物質アデノシンの受容体。カフェインも結合する。
- アドレナリン受容体
- アドレナリンやその他の構造が類似したホルモン、薬物を結合する。
- GABA受容体 (B型)
- アンギオテンシン受容体
- アンギオテンシンの受容体
- カンナビノイド受容体
- 大麻成分およびアナンダミド等の内在性リガンドを結合する。
- コレシストキニン受容体
- コレシストキニンの受容体
- ドーパミン受容体
- ドーパミンの受容体
- オレキシン受容体
- オレキシンの受容体
- グルカゴン受容体
- グルカゴンの受容体
- ヒスタミン受容体
- ヒスタミンの受容体
- 嗅覚受容体
- 嗅覚細胞にある、におい物質の受容体。(2004年度ノーベル生理学・医学賞対象)
- オピオイド受容体
- アヘン成分および内在性ペプチド性リガンド(エンケファリン、エンドルフィン等)を結合する。
- ロドプシン
- 網膜にある光受容体。
- セクレチン受容体
- セクレチンの受容体
- セロトニン受容体
- セロトニンの受容体(3型を除く)
- ソマトスタチン受容体
- ソマトスタチンの受容体
- ガストリン受容体
- ガストリンの受容体
- P2Y受容体
- ATPなどプリンヌクレオチドの受容体
構造

Gタンパク質共役受容体は膜タンパク質 であり、7本の膜貫通ヘリックスを持っている。 細胞外部分はグリコシル化(糖鎖修飾)されていることもある。細胞外ループには2つのよく保存されたシステイン残基が含まれ、ジスルフィド結合によって受容体構造を安定化している。7回貫通型ヘリックスを持つ似たタンパク質には、イオンチャネルとして働くチャネルロドプシンのように、Gタンパク質共役受容体とはまったく異なる機能を持つものがある。
Gタンパク質共役受容体の構造の初期のモデルは、バクテリオロドプシン との弱い相同性に基づいていた。バクテリオロドプシンの構造は、1900年代に電子線回折(PDB: 2BRD, 1AT9)[14][15]や X線結晶構造解析 (1AP9)[16]で解かれていた。 2000年に、哺乳類のGタンパク質共役受容体の構造がウシの ロドプシン (1F88) で初めて解明された[4]。7つの膜貫通ヘリックスなど大まかな構造はバクテリオロドプシンと似ていたが、ヘリックス同士の位置関係は大きく異なっていた。 2007年に、β2-アドレナリン受容体の構造が解かれ、ヒトのGタンパク質共役受容体の構造が初めて明らかになった(2R4R, 2R4S)[5]。この構造は、受容体に抗体を結合して結晶化を補助する方法で得られた。 続いて、受容体の第三細胞内ループを T4 リゾチームで置換した変異体を脂質キュービック相の中で結晶化することにより、より高解像度の構造が得られた(2RH1)[17][18]。 同様の手法で、ドーパミンD3受容体[19]とCXCケモカイン受容体CXCR4[20]の構造も解かれた。 また、シチメンチョウのβ1受容体の熱安定化変異体の構造[21]も報告された。これらのGタンパク質共役受容体の構造は、7本の膜貫通ヘリックス部分ではウシのロドプシンとよく似ていた。しかし、ヘリックス同士を結ぶループ領域の構造はそれぞれ異なっていた。
上記の構造はすべて、アンタゴニストや逆アゴニストが結合した非活性型コンフォメーションである。2011年になって、アゴニストが結合した活性型コンフォメーションと思われる構造が報告された。これらについては後述する。
Remove ads
周辺部との関係
Gタンパク質共役受容体はN末端が細胞外に、C末端が細胞内にあり、疎水性の[22]αヘリックスからなる膜貫通(TM)ドメインが7カ所ある。細胞膜の脂質二重層の部分を7回の膜貫通している。
一方、Gタンパク質共役受容体と共役しているGタンパク質はα(約40 kDa)、β(約35 kDa)、γ(約7〜8 kDa)の3つのサブユニットから構成されている。生理的環境においては、β及びγは互いに固く結合しており、Gβγ複合体と呼ばれる。一方、GαにはGTP/GDP結合部位が存在しており、ここにGDPが結合しているとき、Gタンパク質は不活性型として三量体構造Gαβγを取って、Gタンパク質共役受容体と結合している。2011年になってβ2アドレナリン受容体とGタンパク質の複合体の立体構造が解かれた[23]。
Gタンパク質共役受容体の画像
 アドレナリンβ2受容体の構造(カラゾロールとの複合体)
アドレナリンβ2受容体の構造(カラゾロールとの複合体) アデノシンA2A受容体の構造
アデノシンA2A受容体の構造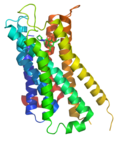 ドーパミンD3受容体の構造(エチクロプリドとの複合体)
ドーパミンD3受容体の構造(エチクロプリドとの複合体) ケモカイン受容体の1種であるCXCR4の構造(IT1tとの複合体)
ケモカイン受容体の1種であるCXCR4の構造(IT1tとの複合体)


.png)
.png)
脚注
関連人物
外部リンク
Wikiwand - on
Seamless Wikipedia browsing. On steroids.
Remove ads