Eurypterida
ordre éteints de chélicèrates De Wikipédia, l'encyclopédie libre

Euryptérides, Scorpions de mer, gigantostracés
Eurypterida

Euryptérides, par Ernst Haeckel
266 collections
| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Chelicerata |
| Clade | Dekatriata |
Les euryptérides (Eurypterida, « aux larges appendices »), ou gigantostracés (« géants caparaçonnés »), forment un ordre fossile d'arthropodes chélicérates fossiles. Marins, ils sont couramment appelés scorpions de mer jusqu'au Carbonifère, lorsqu'une partie d'entre eux deviennent terrestres comme Megarachne, ainsi nommé parce que son céphalothorax ressemble à une araignée de la taille d'une tête humaine. Ils ont vécu de l'Ordovicien moyen au Permien et certaines formes dépassaient 2 mètres de long. Ces grands prédateurs étaient situés en haut des réseaux alimentaires marins de l'ère Paléozoïque.
Description et caractéristiques
Leur allure générale était proche de celle des scorpions actuels : un corps allongé et prolongé par une longue queue articulée, et un céphalothorax équipé de plusieurs paires d'yeux, de pinces préhensiles, mais aussi parfois de râteaux permettant de débusquer des proies dans les sables, de pattes marcheuses et une paire de palettes natatoires (sauf chez les espèces purement benthiques ou devenues terrestres). Ils pouvaient sans doute être des prédateurs efficaces, mais aussi des espèces charognardes ou omnivores comme les actuelles limules. La plupart des espèces présentent un aiguillon effilé au bout de leur queue : comme les scorpions modernes, ils injectaient peut-être un venin à leurs assaillants ou à leurs proies.
Les euryptérides disputent au genre Arthropleura (un myriapode qui mesure jusqu'à 2 m de long) le titre de plus grands arthropode ayant existé sur Terre[1].
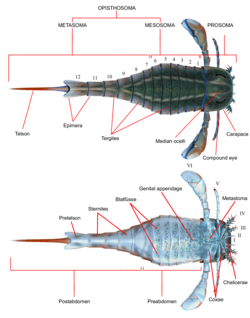 Anatomie d'un euryptéride
Anatomie d'un euryptéride Exemple de fossile bien conservé
Exemple de fossile bien conservé Reconstitution paléoartistique
Reconstitution paléoartistique Modèle 3D
Modèle 3D




Origine, évolution et disparition
Résumé
Contexte
II y a environ 400 millions d'années, les scorpions actuels (qui font partie des arachnides) se séparèrent des euryptérides, en colonisant la terre. Les "scorpions de mer" ne sont donc pas les ancêtres des scorpions actuels, mais plutôt des cousins éloignés. Il est possible que les euryptérides aient mené un mode de vie amphibie en s'accouplant et en pondant en dehors de l'eau grâce à une sorte de deuxième organe respiratoire nommé le "kiemenplatten"[2]. Ils restaient néanmoins dépendants de l'eau et cette dépendance se montrera fatale à leur ordre.
Origine
Le plus ancien euryptéride connu à ce jour a été découvert en 2015. C'est Pentecopterus Decorahensis[3], découvert en Iowa dans des roches datant de 467 millions d'années. Il avait des yeux composés, un corps en deux parties et six paires d'appendices biramés. Une de ces paires lui permettait de se nourrir tandis que les cinq autres lui permettaient de se déplacer.
Au fil du temps, les euryptérides ont commencé à utiliser leurs appendices de deux façons différentes. Ils se sont scindés ainsi en deux sous-ordres distincts[4] : les stylonurinés qui utilisaient leurs appendices pour ramper sur le fond de l'océan et les euryptérinés dont une des paires d'appendices s'était transformée en une sorte de pagaie qui leur permettait de nager.

Le premier stylonuriné connu est Brachyopterus. Il mesurait moins de 20 cm de long. Comme les autres membres de son sous-ordre, pour éviter la concurrence des poissons qui connaissaient un fort développement, il se nourrissait d'animaux morts qui tombaient sur le fond marin.
À la fin de l'Ordovicien, un deuxième sous-ordre apparait dans les fossiles : celui des euryptérinés. Dès leur apparition, ils sont entrés en compétition directe avec les poissons sans mâchoires et avec les céphalopodes. La mobilité de ce groupe leur a permis de se diversifier rapidement tout au long du silurien. En effet, ils ont pu coloniser de nouveaux espaces et s'y développer[4].
Diversification

Du fait de leur compétition avec les poissons et les céphalopodes, ils ont été soumis à une pression sélective et ont dû évoluer pour rivaliser avec les autres prédateurs marins. Ce sous-ordre s'est considérablement diversifié et développé, ce qui explique pourquoi 75% de tous les scorpions de mers y appartiennent. Mais, avec le début du Dévonien, de nouveaux concurrents firent leur apparition : les placodermes, ces grands poissons couverts de plaques osseuses.
Après avoir connu un pic de diversité au tout début du Dévonien[4], certaines espèces se sont éteintes car elles ne pouvaient plus rivaliser avec ces nouveaux prédateurs. Les survivants eux commencèrent une véritable course évolutive pour être plus grands, plus forts et plus rapides. C'est la famille des pterygotidés qui a survécu près de 40 millions d'années de plus.
Les pterygotidés[4]
Sillonnant les mers sans relâche, ils étaient de redoutables prédateurs avec des exosquelettes fins et légers, leur offrant une grande rapidité, des queues aplaties (nommées telson[5]) leur offrant une puissance de nage supplémentaire, une vue perçante et des sortes de pinces nommé chélicères se terminant par des crochets robustes.

La plupart des membres de cette famille dépassaient le mètre de long, mais le plus grand d'entre eux, le Jaekelopterus mesurait 2,5 mètres. Sa pince mesurait 50 cm de long et ses crochets avaient la taille d'une canine de tigre[6].
Mais, malgré leurs efforts, ils ne pouvaient pas rivaliser avec les placodermes et ils perdirent 50% de leur diversité au cours des 10 premiers millions d'années du Dévonien.
Survie et déclin des stylonurinés

À la fin du Dévonien, tous les eurypterinés avaient disparu. Mais les stylonurinés qui rampaient au fond de l'océan survécurent[4]. En effet, ils étaient moins influencés par l'arrivée des placodermes, d'autant plus qu'ils colonisaient des écosystèmes d'eau douce où il y avait considérablement moins de poissons.
Avec la formation de la Pangée, certains colonisèrent le super-continent tandis que d'autres restèrent dans les eaux douces, comme par exemple, l'hibbertopterus, considéré comme le dernier grand scorpion de mer.
Mais eux aussi finirent par s'éteindre à la fin du Permien, à la suite de la plus grande extinction de l'histoire de la Terre. Ainsi s'acheva l'ère des euryptérides qui dura plus de 210 millions d'années.
Taille
Résumé
Contexte

La taille des euryptérides était très variable et dépendait de facteurs tels que le mode de vie, l'environnement et les affinités taxonomiques. Des tailles d'environ 1 mètre sont courantes dans la plupart des groupes[7]. Le plus petit euryptéride, Alkenopterus burglahrensis, ne mesurait que 2,03 centimètres de long[8].
Le plus grand euryptéride, et le plus grand arthropode connu, est Jaekelopterus rhenaniae. Deux autres euryptérides pourraient également avoir atteint une longueur de 2,5 mètres : Erettopterus grandis (étroitement apparenté à Jaekelopterus) et Hibbertopterus wittebergensis, mais E. grandis est très fragmentaire et l'estimation de la taille de H. wittenbergensis est basée sur des traces de déplacement sur le fond, et non sur des restes fossilisés[9].
La famille des Jaekelopterus : les Pterygotidae, est connue pour plusieurs espèces de grande taille. Acutiramus, dont le plus grand membre, A. bohemicus, mesurait 2,1 mètres, et Pterygotus, dont la plus grande espèce, P. grandidentatus, mesurait 1,75 mètre, en font partie[10]. Plusieurs facteurs différents ont été suggérés pour expliquer la grande taille des ptérygotides, notamment la séduction, la facilité de prédation et la situation aux sommets des réseaux trophiques[11].
Les grands euryptérides ne se trouvaient pas seulement parmi les Pterygotidae : un métasome fossile isolé de 12,7 centimètres de long de l'euryptéride carcinosomatoïde Carcinosoma punctatum indique que l'animal aurait atteint une longueur de 2,2 mètres, rivalisant en taille avec les ptérygotides[12]. Un autre géant était Pentecopterus decorahensis, un carcinosomatoïde primitif, qui aurait atteint une longueur de 1,7 mètre[13]. D'une manière générale, les arthropodes sont limités en taille par leur système de respiration et des facteurs tels que la locomotion, les coûts énergétiques de la mue et les propriétés physiques de l'exosquelette, mais les paléontologues pensent que des branchies externes de grande taille (parties souples non fossilisées car hors de l'exosquelette) ont pu faciliter leur oxygénation, permettant à quelques euryptérides de se développer grâce à un air et donc à une eau qui étaient plus riches en oxygène (jusqu'à 35 %) qu'aujourd'hui (21 %)[14],[15].
Les euryptérides de grande taille sont généralement de constitution légère, la plupart des grands segments corporels fossilisés étant minces et non minéralisés. Ces adaptations permettaient d’alléger les exosquelettes et sont également présentes chez d'autres arthropodes géants du Paléozoïque, comme Arthropleura, un mille-pattes de 2 m de long[10],[16].
Registre fossile
L'espèce connue la plus ancienne d'euryptéride est Pentecopterus decorahensis découverte en 2015, et datant du début du Darriwilien (Ordovicien moyen). Cette espèce était déjà relativement complexe, ce qui suggère que les premiers euryptérides seraient apparus dès le début de l'Ordovicien, il y a environ 485 millions d'années[1].
Les euryptérides connurent un grand succès évolutif au Silurien et au Dévonien où ils étaient parmi les principaux super-prédateurs, mais seuls deux groupes survécurent à la fin du Dévonien (les Adelophtalmoidea et les Stylonurina), qui disparurent à leur tour au Permien. Leur succès évolutif a donc duré 250 millions d'années entre environ -500 et -250 millions d'années avant le présent, soit autant de temps qu'il s'en est écoulé depuis leur disparition[1].
Classification
Résumé
Contexte
Systématique des taxons d'euryptérides sur le modèle de Dunlop et al. (2023)[17]. Les taxons et leurs auteurs sont principalement issus de ce travail, avec quelques additions d'autres sources en cas d'absence de taxons ou d'auteurs[17],[18],[19],[20],[21],[22]. Les Megalograptidae sont considérés comme des Carcinosomatoidea d'après plusieurs travaux récents[20],[23].
Sous-ordre Stylonurina Diener, 1924:
- Super-famille Rhenopteroidea Størmer, 1951
- Famille Rhenopteridae Størmer, 1951
- Brachyopterella Kjellesvig-Waering, 1966
- Brachyopterus Størmer, 1951
- Kiaeropterus Waterston, 1979
- Leiopterella Lamsdell et al., 2010
- Rhenopterus Størmer, 1936
- Famille Rhenopteridae Størmer, 1951
- Super-famille Stylonuroidea Kjellesvig-Waering, 1959
- Famille Parastylonuridae Waterston, 1979
- Parastylonurus Kjellesvig-Waering, 1966
- Stylonurella Kjellesvig-Waering, 1966
- Famille Stylonuridae Diener, 1924
- Ctenopterus Clarke & Ruedemann, 1912
- Laurieipterus Kjellesvig-Waering, 1966
- Pagea Waterston, 1962
- Soligorskopterus Plax et al., 2018
- Stylonurus Page, 1856
- Famille Parastylonuridae Waterston, 1979
- Super-famille Kokomopteroidea Kjellesvig-Waering, 1966
- Famille Kokomopteridae Kjellesvig-Waering, 1966
- Kokomopterus Kjellesvig-Waering, 1966
- Lamontopterus Waterston, 1979
- Famille Hardieopteridae Tollerton, 1989
- Hardieopterus Waterston, 1979
- Hallipterus Kjellesvig-Waering, 1963
- Tarsopterella Størmer, 1951
- Famille Kokomopteridae Kjellesvig-Waering, 1966
- Super-famille Mycteropoidea Cope, 1886
- Famille Drepanopteridae Kjellesvig-Waering, 1966
- Drepanopterus Laurie, 1892
- Famille Hibbertopteridae Kjellesvig-Waering, 1959
- Campylocephalus Eichwald, 1860
- Cyrtoctenus Størmer & Waterston, 1968
- Dunsopterus Waterston, 1968
- Hastimima White, 1908
- Hibbertopterus Kjellesvig-Waering, 1959
- Vernonopterus Waterston, 1957
- Famille Mycteropidae Cope, 1886
- Megarachne Hünicken, 1980
- Mycterops Cope, 1886
- Woodwardopterus Kjellesvig-Waering, 1959
- Famille Drepanopteridae Kjellesvig-Waering, 1966
- Incertae sedis
- Stylonuroides Kjellesvig-Waering, 1966
Sous-ordre Eurypterina Burmeister, 1843:
- Super-famille Moselopteroidea Lamsdell et al., 2010
- Famille Moselopteridae Lamsdell et al., 2010
- Moselopterus Størmer, 1974
- Stoermopterus Lamsdell, 2011
- Vinetopterus Poschmann & Tetlie, 2004
- Famille Moselopteridae Lamsdell et al., 2010
- Super-famille Onychopterelloidea Lamsdell, 2011
- Famille Onychopterellidae Lamsdell 2011
- Alkenopterus Størmer, 1974
- Onychopterella Størmer, 1951
- Tylopterella Størmer, 1951
- Famille Onychopterellidae Lamsdell 2011
- Super-famille Eurypteroidea Burmeister, 1843
- Famille Erieopteridae Tollerton, 1989
- Erieopterus Kjellesvig-Waering, 1958
- Famille Eurypteridae Burmeister, 1843
- Eurypterus De Kay, 1825
- Famille Dolichopteridae Kjellesvig-Waering and Stromer, 1952
- Clarkeipterus Kjellesvig-Waering, 1966
- Dolichopterus Hall, 1859
- Malongia Wang et al., 2022
- Ruedemannipterus Kjellesvig-Waering, 1966
- Famille Strobilopteridae Lamsdell & Selden, 2013
- Buffalopterus Kjellesvig-Waering & Heubusch, 1962
- Strobilopterus Ruedemann, 1935
- Incertae sedis
- Paraeurypterus Hoşgör & Selden, 2013
- Pentlandopterus Hoşgör & Selden, 2013
- Famille Erieopteridae Tollerton, 1989
- Infra-ordre Diploperculata Hoşgör & Selden, 2013
- Super-famille Carcinosomatoidea Størmer, 1934
- Famille Megalograptidae Caster & Kjellesvig-Waering, 1955
- Echinognathus Walcott, 1882
- Megalograptus Miller, 1874
- Pentecopterus Lamsdell et al., 2015
- Famille Carcinosomatidae Størmer, 1934
- Carcinosoma Claypole, 1890
- Eocarcinosoma Caster & Kjellesvig-Waering, 1964
- Paracarcinosoma Caster & Kjellesvig-Waering, 1964
- Rhinocarcinosoma Novojilov, 1962
- Famille Mixopteridae Caster & Kjellesvig-Waering, 1955
- Lanarkopterus Ritchie, 1968
- Mixopterus Ruedemann, 1921
- Terropterus Wang et al., 2021
- Famille Megalograptidae Caster & Kjellesvig-Waering, 1955
- Super-famille Waeringopteroidea Tetlie, 2004 (super-famille dérivée d'une thèse non-publiée)
- Orcanopterus Stott et al., 2005
- Grossopterus Størmer, 1934
- Waeringopterus Leutze, 1961
- Super-famille Adelophthalmoidea Tollerton, 1989
- Famille Adelophthalmidae Tollerton, 1989
- Adelophthalmus Jordan in Jordan & von Mayer, 1854
- Archopterus Wang et al., 2023
- Bassipterus Kjellesvig-Waering & Leutze, 1966
- Eysyslopterus Tetlie & Poschmann, 2008
- Nanahughmilleria Kjellesvig-Waering, 1961
- Parahughmilleria Kjellesvig-Waering, 1961
- Pittsfordipterus Kjellesvig-Waering & Leutze, 1966
- Pruemopterus Poschmann, 2021
- Famille Adelophthalmidae Tollerton, 1989
- Super-famille Pterygotioidea Clarke & Ruedemann, 1912
- Famille Hughmilleriidae Kjellesvig-Waering, 1951
- Herefordopterus Tetlie, 2006
- Hughmilleria Sarle, 1903
- Famille Slimoniidae Novojilov, 1968
- Salteropterus Kjellesvig-Waering, 1951
- Slimonia Page, 1856
- Famille Pterygotidae Clarke & Ruedemann, 1912
- Acutiramus Ruedemann, 1935
- Ciurcopterus Tetlie & Briggs, 2009
- Erettopterus Salter in Huxley & Salter, 1859
- Jaekelopterus Waterston, 1964
- Necrogammarus Woddward, 1870
- Pterygotus Agassiz, 1839
- Famille Hughmilleriidae Kjellesvig-Waering, 1951
- Super-famille Carcinosomatoidea Størmer, 1934
- Incertae sedis
- Dorfopterus Kjellesvig-Waering, 1955
- Holmipterus Kjellesvig-Waering, 1979
- Marsupipterus Caster & Kjellesvig-Waering, 1955
- Unionopterus Chernyshev, 1948
 Rhenopterus diensti (Rhenopteridae)
Rhenopterus diensti (Rhenopteridae) Stylonurella spinipes (Parastylonuridae)
Stylonurella spinipes (Parastylonuridae)

 Kokomopterus longicaudatus (Kokomopteridae)
Kokomopterus longicaudatus (Kokomopteridae) Hardieopterus sp. (Hardieopteridae)
Hardieopterus sp. (Hardieopteridae)


 Stoermeropterus sp. (Moselopteridae)
Stoermeropterus sp. (Moselopteridae) Onychopterella augusti (Onychopterellidae)
Onychopterella augusti (Onychopterellidae) Erieopterus sp. (Erieopteridae)
Erieopterus sp. (Erieopteridae) Eurypterus sp. (Eurypteridae), vue dorsale et ventrale
Eurypterus sp. (Eurypteridae), vue dorsale et ventrale Dolichopterus macrocheirus (Dolichopteridae)
Dolichopterus macrocheirus (Dolichopteridae) Strobilopterus proteus (Strobilopteridae)
Strobilopterus proteus (Strobilopteridae)
 Carcinosoma newlini (Carcinosomatidae)
Carcinosoma newlini (Carcinosomatidae) Mixopterus kiaeri (Mixopteridae)
Mixopterus kiaeri (Mixopteridae) Waeringopterus sp. (Waeringopteroidea)
Waeringopterus sp. (Waeringopteroidea) Adelophthalmus irinae (Adelophthalmidae)
Adelophthalmus irinae (Adelophthalmidae) Hughmilleria socialis (Hughmilleriidae)
Hughmilleria socialis (Hughmilleriidae) Slimonia acuminata (Slimoniidae)
Slimonia acuminata (Slimoniidae)



_(14596592278).jpg)







.jpg)
.png)





.jpg)




Phylogénie
Place au sein des chélicérates
Phylogénie des grands groupes de chélicérés, d'après Lamsdell, 2013[24] :
| Chelicerata |
| ||||||||||||||||||||||||
Voir aussi
Articles connexes
Lien externe
Références taxinomiques
Notes et références
Wikiwand - on
Seamless Wikipedia browsing. On steroids.