
From Wikipedia, the free encyclopedia
U genetici, umrežavanje DNK označava pojave kada različiti egzogeni ili endogeni agensi reaguju sa dva nukleotida DNK, formirajući kovalentnu vezu između njih. Ova poprečna veza može se desiti unutar istog lanca (unutarlančana) ili između suprotnih lanaca dvolančane DNK (međulančana). Ovi adukti ometaju ćelijski metabolizam, kao što su replikacija DNK i transkripcija, pokrećući ćelijsku smrt. Ove poprečne veze se, međutim, mogu popraviti putem ekscizije ili rekombinacijskih puteva.

Umrežavanje DNK također ima korisne zasluge u hemoterapijama i ciljanju kancerogenih ćelija za apoptoze,[1] kao i u razumijevanju interakcije proteina sa DNK.
Mnogi karakterizirani agensi za umrežavanje imaju dvije neovisno reaktivne grupe unutar iste molekule, od kojih se svaka može vezati za nukleotidni ostatak DNK. Ovi agensi su odvojeni na osnovu njihovog izvora i označeni kao egzogeni ili endogeni. Egzogeni agensi za umrežavanje su hemikalije i spojevi, prirodni i sintetski, koji proizlaze iz izloženosti okolišu kao što su lijekovi i dim cigareta ili izduvni gasovi automobila. Endogeni agensi za umrežavanje su spojevi i metaboliti koji se unose iz ćelijskih ili biohemijskih puteva unutar ćelije ili organizma.
| Umrežavajući agens | Alkilirajući agens | Struktura umreživača | Preferencijalna ciljna sekvenca |
| Dušični senf |   |
Unujtarlančano; 5'-GC | |
| Cisplatin |  |
 |
Unutarlančano; 5'-GC
Međulančano; 5'-GNG |
| Karmustin (BNCU) |  |
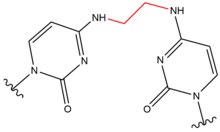 |
Međulančano; bazni par GC |
| Mitomicin C |  |
 |
Međulančano; 5'-GC |
| Psoralen |   |
Međulančano; 5'-TA | |
| Malondialdehid |  |
 |
Međulančano; 5'-GC, 5'-CG |
| Oksidativni stres (i zračrnje) |  |
Unutarlančano; d(GpT) | |
| Dušična kiselina |  |
 |
Međulančano; 5'-CG |
Umrežena DNK se popravlja u ćelijama kombinacijom enzima i drugih faktora iz puta nukleotidnog ekscizijskog popravka (NER), homolognom rekombinacijom i puta baznoekscijskog popravka (BER). Za popravak međulančanih umreženih veza kod eukariota, endonukleaza od 3' preklopa iz NER-a, XPF-ERCC1, regrutuje se u umreženu DNK, gdje pomaže u 'otkačivanju' DNK, cijepanjem 3' lanca na mjestu umrežavanja. 5' lanac se zatim cijepa, bilo pomoću XPF-ERCC1 ili druge endonukleaze, formirajući dvolančani prekid (DSB), koji se zatim može popraviti pomoću puta homologne rekombinacije.[17]
Umređena DNK općenito uzrokuju gubitak informacija o preklapanju sekvenci iz dva lanca DNK. Stoga, tačna popravka oštećenja ovisi o preuzimanju izgubljene informacije iz neoštećenog homolognog hromosoma u istoj ćeliji. Preuzimanje se može dogoditi uparivanjem sa sestrinskom hromatidom proizvedenom tokom prethodnog kruga replikacije. U diploidnoj ćeliji umrežavanje se takođe može desiti uparivanjem sa nesestrinskim homolognim hromosomom, što se dešava posebno tokom mejoze. Kada dođe do uparivanja, umrežavanje se može ukloniti i ispravne informacije uvesti u oštećeni hromosom, homolognom rekombinacijom.
Raskid veze između šećera dezoksiriboze u šećerno-fosfatnoj kičmi DNK i povezane nukleobaza ostavlja abazno mjesto u dvolančanoj DNK. Ova abazna mjesta se često stvaraju kao međuproizvodi, a zatim se obnavljaju u baznoj ekscizijskoj popravci. Međutim, ako se ovim mjestima dopusti da opstanu, mogu inhibirati replikaciju i transkripciju DNK.[18] Abazna mjesta mogu reagovati sa aminskim grupama na proteinima da bi formirali umrežavanje DNK-protein ili sa egzocikličkim aminima drugih nukleobaza da bi formirale međulančane umrežene veze. Kako bi spriječili međulančane ili umrežavanje DNK-protein, enzimi iz BER puta čvrsto vezuju abazno mjesto i izdvajaju ga iz obližnjih reaktivnih grupa, kao što je pokazano u ljudskoj alkiladenin DNK glikozilazi (AAG) i E. coli 3-metiladenin DNK glikozilazi II (AlkA).[19] In vitro dokazi pokazali su da međulančano umrežavanje inducirano nebaznim mjestom (DOB-ICL) predstavljaju lezije koje blokiraju replikaciju i vrlo pogrešno kodiraju. U poređenju sa nekoliko drugih ispitanih TLS pol-a, pol η će vjerovatno doprinijeti TLS-posredovanoj popravci DOB-ICL in vivo.[20] Korištenjem O6-2'-deoksigvanozin-butilen-O6-2'-deoksiguanozin (O6-dG-C4-O6-dG) lezija DNK koja je hemijski stabilna struktura, istraživana je aktivnost premošćavanja nekoliko DNK polimeraza i rezultati su pokazali da pol η ima najveću aktivnost premošćavanja; međutim, 70% bajpas proizvoda je bilo mutageno i sadrži zamjene ili delecije. Povećanje veličine nepovezanih popravnih intermedijera podiže učestalost delecijskih mutacija.[21]
Tretiranje E. coli sa psoralenom-plus-UV svjetlom (PUVA) stvara međulančano umredžavanje DNK. Cole et al.[22] i Sinden & Cole[23] predstavili su dokaz da homolognorekombinacijski proces popravke koji zahtijeva proizvode gena uvrA, uvrB i recA može ukloniti ova umrežavanja u E. coli. Čini se da je ovaj proces prilično efikasan. Iako su jedno ili dva nepopravljena umrećavanja dovoljna da inaktiviraju ćeliju, bakterijska ćelija divljeg tipa može se popraviti i stoga oporaviti od 53 do 71 psoralenskih umrežavanja. Eukariotske ćelije kvasca su također inaktivirane jednom preostalom umreženom vezom, ali ćelije kvasca divljeg tipa mogu se oporaviti od 120 do 200 umrežavanja.[24]
Umrežavanje DNK-protein može biti uzrokovano raznim hemijskim i fizičkim agensima, uključujući prijelazne metale, ionizirajuće zračenje i endogene aldehide, pored hemoterapeutskih agenasa.[25] eDNK umrežavanju, DNK-protein umrežavanju su lezije u ćelijama koje se često oštećuju UV zračenjem. Utjecaj UV zraka može dovesti do reaktivnih interakcija i uzrokovati umrežavanje DNK i proteina koji su u kontaktu s njom. Ove umrežavajuće veze su veoma glomazne i složene lezije. One se prvenstveno javljaju u područjima hromosoma koji prolaze kroz replikaciju DNK i ometaju ćelijske procese.
Napredak u metodima identifikacije strukture je napredovao, a dodatak sposobnosti mjerenja interakcija između DNK i proteina je zahtjev za potpuno razumijevanje biohemijskih procesa. Struktura kompleksa DNK-protein može se mapirati pomoću fotoumrežavanja, što je fotoindukovano formiranje kovalentne veze između dva makromolekula ili između dva različita dijela jedne makromolekule. Metodologija uključuje kovalentno povezivanje DNK-vezujućeg motiva DNK-vezujućeg proteina specifičnog za ciljnu sekvencu sa agensom za umrežavanje koji se može fotoaktivirati i koji je sposoban da reaguje sa nukleotidima DNK kada je izložen UV zračenju. Ovaj metoda daje informacije o interakciji između DNK i proteina u unakrsnoj vezi umrežavanja.[26]
Putevi popravka DNK mogu dovesti do stvaranja tumorskih ćelija. Kancerski tretmani su dizajnirani pomoću agenasa za umrežavanje DNK za interakciju sa dušičnim bazama DNK kako bi se blokirala replikacija DNK. Ovi agensi za umrežavanje imaju sposobnost da djeluju kao terapije sa jednim agensom ciljajući i uništavajući specifične nukleotide u ćelijama raka. Rezultat je zaustavljanje ciklusa i rasta ćelija raka; budući da inhibira specifične puteve popravka DNK, ovaj pristup ima potencijalnu prednost jer ima manje nuspojava.[27]
Kod ljudi, vodeći uzrok smrti od raka u svijetu je rak pluća, uključujući karcinom nemalih a plućnih ćelija (NSCLC) koji čini 85% svih slučajeva raka pluća u Sjedinjenim Državama.[28] Pacijenti sa NSCLC liječe se često terapeutskim spojevima platine (npr. cisplatin, karboplatin ili oksaliplatin) koji uzrokuju međulančano umrežavanje DNK. Među osobama sa NSLC-om, niska ekspresija gena raka dojke 1 (BRCA1) u primarnom tumoru korelirala je s poboljšanim preživljavanjem nakon hemoterapije koja sadrži platinu.[29][30] Ova korelacija implicira da nizak BRCA1 u kanceru, i posljedično nizak nivo popravka DNK, uzrokuje ranjivost raka na liječenje agensima za umrežavanje DNK. Visok BRCA1 može zaštititi ćelije raka djelujući u homolognorekombinacijskom putu popravka koji uklanja oštećenja u DNK unesena lijekovima platine. Nivo ekspresije BRCA1 potencijalno je važan alat za prilagođavanje hemoterapije u liječenju raka pluća.[29][30]
Klinički hemoterapeutici mogu inducirati enzimsko i neenzimsko umrežavanje DNK-protein. Primjer ove indukcije su derivati platine, kao što su cisplatin i oksaliplatin. Oni stvaraju neenzimsko umrežavanje DNK-protein kroz nespecifično umrežavanje proteina koji stupaju u interakciju s hromatinom za DNK. Umrežavanje je moguće i kod drugih terapeutskih agenasa, bilo stabilizacijom intermedijara reakcije kovalentne DNK-protein ili stvaranjem pseudosupstrata, koji hvata enzim na DNK. Derivati kamptotecina, kao što su irinotekan i topotekan, ciljaju i hvataju specifičnu DNK topoizomerazu 1 (TOP1), interkalacijom unutar interfejsa enzim-DNK. Budući da toksičnost ovih lijekova ovisi o hvatanju TOP1, ćelijska osjetljivost na ove spojeve direktno zavisi od nivoa ekspresije TOP1. Kao rezultat toga, funkcija ovih lijekova je da služe kao otrovi enzima, a ne kao inhibitori. Ovo se može primijeniti za liječenje tumorskih ćelija, korištenjem TOP 2 otrova enzima.[31]
Seamless Wikipedia browsing. On steroids.
Every time you click a link to Wikipedia, Wiktionary or Wikiquote in your browser's search results, it will show the modern Wikiwand interface.
Wikiwand extension is a five stars, simple, with minimum permission required to keep your browsing private, safe and transparent.