阿斯加德古菌[2]是古菌的一個類群,屬於提議中的阿斯加德古菌超門[3][4](Asgard或Asgardarchaeota[5]),包含了洛基古菌門、索爾古菌門、奧丁古菌門跟海姆達爾古菌門等。[6]研究顯示真核生物似乎演化自一類屬於阿斯加德古菌超門、包含海姆達爾古菌門的生物,[7][8]而這為兩域系統而非三域系統提供了支持。[9][10]
Quick Facts 阿斯加德古菌, 科學分類 ...
阿斯加德古菌
|
科學分類 
|
| 域:
|
古菌域 Archaea
|
| 界:
|
變形古菌界 Proteoarchaeota
|
| 總門:
|
阿斯加德古菌超門 Asgard
Katarzyna Zaremba-Niedzwiedzka(), et al. 2017
|
| 門
|
- 洛基古菌門 Lokiarchaeota
- 索爾古菌門 Thorarchaeota
- 奧丁古菌門 Odinarchaeota
- 海姆達爾古菌門 Heimdallarchaeota
|
|
| 異名
|
- "Asgardarchaeota" Violette Da Cunha et al. 2017
- "Asgardaeota" Whitman 2018
- "Eukaryomorpha" Fournier & Poole 2018[1]
|
Close
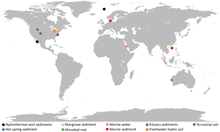
2010年夏天,從北冰洋科尼波維奇海嶺(Knipovich ridge)的裂谷一個名為洛基城堡熱泉的海底熱泉附近取得的沉積物岩芯樣本被送交分析,其中先前發現有豐富新古菌物種的沉積物水平面受到了宏基因組分析。
2015年,一組由瑞典烏普薩拉大學領導的團隊在使用高度保守的蛋白質基因序列對這些生物進行系統發生的分析後,提出了洛基古菌門的分類[11],而其名稱來自北歐神話中的變形神洛基,這名稱同時也取自做為這些樣本來源的海底熱泉[12];而北歐神話的洛基被認為是一個「驚人地複雜、令人困惑,且模稜兩可的人物,而他也是無數未解決的學術爭論中的催化劑。」[13]而這也與洛基古菌門在關於真核生物起源的爭辯中所扮演的角色相呼應。[11][14]
2016年,一組由美國德州大學領導的團隊從源自美國北卡羅萊納州白橡河的樣本中發現了索爾古菌門,而這類生物的名稱源自另一個北歐神話的神祉索爾。[15]
此外,更多來自洛基城堡熱泉、美國黃石國家公園、丹麥奧胡斯灣、美國科羅拉多河附近的含水層、紐西蘭輻射池、日本竹富島附近海底熱泉以及美國白橡河河口的等地的樣本,使得學者隨後發現了奧丁古菌門跟海姆達爾古菌門等其他的生物;[6]而由於已約定俗成說這類生物以北歐神話的人物命名之故,因此這些古菌也分別以奧丁跟海姆達爾等北歐神話人物命名,也因此研究人員將這些微生物組成的超門給依照北歐神話中諸神所住的領域「阿斯加德」給命名為「Asgard」。[6]
阿斯加德古菌具有很多蛋白質與真核生物中相對應的蛋白質相似,這其中包括了鳥苷三磷酸酶、ESCRT及CHMP4B等細胞膜蛋白質、泛素修飾系統,以及N-糖基化途徑同源基因等。[6]
阿斯加德古菌有受調控的肌動蛋白細胞骨架,且其所用的前纖維蛋白和凝溶膠蛋白可與真核生物的肌動蛋白產生作用;[16][17]此外,源自棲息於熱液之中的奧丁古菌的微管蛋白,也就是奧丁微管蛋白(OdinTubulin),被認為是真正的微管蛋白;奧丁微管蛋白所構成的原體和原纖絲與真核生物的微管最為接近,但也會組成一個類似FtsZ的環狀系統,這表示奧丁微管蛋白可能是FtsZ和構成微管的微管蛋白之間的演化環節;[18]不僅如此,在低溫電子顯微鏡的觀察下,這些生物似乎有囊泡,一些生物可能還有屬於PKD域的S層蛋白;[19]此外,這些生物的核糖體大亞基也與真核生物一般,有三向ES39擴張。[20]
-
阿斯加德古菌的代謝途徑,因生物所屬的門而異
[21]
-
阿斯加德古菌一般是專性厭氧菌;然而卡瑞古菌門(Kariarchaeota)、葛德古菌門(Gerdarchaeota)與霍德爾古菌門(Hodarchaeota)可能是兼性厭氧菌。[22]
這些生物使用伍德-隆達爾代謝途徑並進行糖酵解;這門的生物可能是自營生物、異營生物或者使用太陽視紫紅質的光營生物;[21]而其中的一種生物Prometheoarchaeum syntrophicum與一種減硫變形菌和一種產甲烷古菌互養。[19]
阿斯加德古菌的RuBisCO不用於固碳,但可能用於核苷酸的補救合成。[21]
在2017年,研究發現海姆達爾古菌有著N-末端核心組蛋白尾(N-terminal core histone tails),而這特徵先前被認為只存在於真核生物上;之後在2018年,學者發現有兩個阿斯加德古菌超門以外的門的古菌也有這樣的組蛋白尾。[23]
在2020年一月,科學家發現說洛基古菌門的生物Prometheoarchaeum syntrophicum與兩種細菌互養,考慮這與共生體學說的相似性,他們認為這種關係與簡單原核微生物與複雜真核微生物在大約二十億年發生的事之間有所關聯。[24][19]
阿斯加德古菌間的系統發生關係尚在討論中。
| Williams et al. 2019,[8] Eme et al. 2017,[7] Liu et al. 2021[25] & Liu et al. 2020[22]
|
GTDB release 07-RS207 (8th April 2022)[26][27][28]
|
| 變形古菌界
|
|
|
TACK超門
|
|
|
| 阿斯加德古菌超門
|
|
|
|
|
包爾古菌門(Borrarchaeota)
|
|
|
|
|
|
|
|
|
赫爾古菌門(Helarchaeota)
|
|
|
|
|
洛基古菌門(Lokiarchaeota)
|
|
|
|
|
|
|
|
|
|
|
|
巴德爾古菌門(Baldrarchaeota)
|
|
|
|
|
奧丁古菌門(Odinarchaeota)
|
|
|
|
|
|
|
|
|
|
赫爾莫德古菌門(Hermodarchaeota)
|
|
|
|
|
索爾古菌門(Thorarchaeota)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
悟空古菌門(Wukongarchaeota)
|
|
|
|
|
|
|
霍德爾古菌門(Hodarchaeota)
|
|
|
|
|
|
|
葛德古菌門(Gerdarchaeota)
|
|
|
|
|
|
|
卡瑞古菌門(Kariarchaeota)
|
|
|
|
|
海姆達爾古菌門(Heimdallarchaeota)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| (+α-變形菌)
|
真核生物
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| 希芙古菌綱
|
|
|
包爾古菌目(Borrarchaeales)
|
|
|
|
|
希芙古菌目(Sifarchaeales)
|
|
|
|
|
|
|
|
| 悟空古菌綱
|
悟空古菌目(Wukongarchaeales)
|
|
|
| (Heimdallarchaeia)
|
|
|
霍德爾古菌目(Hodarchaeales)
|
|
|
|
|
|
|
葛德古菌目(Gerdarchaeales)
|
|
|
|
|
|
|
卡瑞古菌目(Kariarchaeales)
|
|
|
|
|
海姆達爾古菌目(Heimdallarchaeales)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| 嬌德古菌綱
|
嬌德古菌目(Jordarchaeales)
|
|
|
| 奧丁古菌綱(Odinarchaeia)
|
奧丁古菌目(Odinarchaeales)
|
|
|
|
|
|
|
|
|
|
| 巴德爾古菌綱
|
巴德爾古菌目(Baldrarchaeales)
|
|
|
| 索爾古菌綱(Thorarchaeia)
|
索爾古菌目(Thorarchaeales)
|
|
|
|
|
|
|
|
| 赫爾莫德古菌綱
|
赫爾莫德古菌目(Hermodarchaeales)
|
|
|
| 洛基古菌綱(Lokiarchaeia)
|
|
|
赫爾古菌目(Helarchaeales)
|
|
|
|
|
洛基古菌目(Lokiarchaeales)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
在上述的場景中,真核生物位於阿斯加德古菌超門的分類樹的深處,而比起洛基古菌等,真核生物與海姆達爾古菌更為接近,一些人提議說應該把海姆達爾古菌分成多個類群(海姆達爾古菌門、卡瑞古菌門、葛德古菌門與霍德爾古菌門等)。真核生物可能是前述的四個群體與悟空古菌門(Wukongarchaeota)的姊妹群,也有可能是整個阿斯加德古菌超門的姊妹群。目前學界較偏好互養場景,也就是兩類有生物彼此互相滋養的場景,在這種狀況下,真核生物可能是阿斯加德古菌嵌入某種未知的細菌、並因此發展成細胞核所致;而某種嵌入這複合體的α-變形菌則變成粒線體。[29]
以下的分類架構是基於命名中出現的原核生物名稱列表(LPSN)[30]和美國國家生物技術資訊中心(NCBI)的說法而生的,該架構如下:[31]
- 弗雷古菌門(Freyrarchaeota,尚未確定)Caceres 2019 ex Xie et al. 2021
- 弗麗嘉古菌門(Friggarchaeota,尚未確定)Caceres 2019
- 葛馮古菌門(Gefionarchaeota,尚未確定)Caceres 2019
- 伊登古菌門(Idunnarchaeota,尚未確定)Caceres 2019
- 尼約德古菌門(Njordarchaeota,尚未確定)Xie et al. 2021
- 西格恩古菌門(Sigynarchaeota,尚未確定)Xie et al. 2021
- 提爾古菌門(Tyrarchaeota,尚未確定)Xie et al. 2021古菌門()
- 希芙古菌綱(Sifarchaeia)Sun et al. 2021
- 包爾古菌目(Borrarchaeales)Liu et al. 2020
- 包爾古菌科(Borrarchaeaceae)Liu et al. 2020
- (候選分類)包爾古菌屬(Borrarchaeum)Liu et al. 2020
- 希芙古菌目(Sifarchaeales)Sun et al. 2021
- 希芙古菌科(Sifarchaeaceae)Sun et al. 2021
- (候選分類)希芙古菌屬(Sifarchaeum)corrig. Farag, Zhao & Biddle 2020
- 悟空古菌綱(Wukongarchaeia)Liu et al. 2020
- 悟空古菌目(Wukongarchaeales)Liu et al. 2020
- 悟空古菌科(Wukongarchaeaceae)Liu et al. 2020
- (候選分類)悟空古菌屬(Wukongarchaeum)Liu et al. 2020
- 海姆達爾古菌綱(Heimdallarchaeia)
- 葛德古菌目([Gerdarchaeales)(JABLTI01)
- 海姆達爾古菌目(Heimdallarchaeales)
- 霍德爾古菌目(Hodarchaeales)Liu et al. 2020
- 霍德爾古菌科(Hodarchaeaceae)Liu et al. 2020
- (候選分類)霍德爾古菌屬(Hodarchaeum)Liu et al. 2020
- 卡瑞古菌目(Kariarchaeales)Liu et al. 2020
- 卡瑞古菌科(Kariarchaeaceae)Liu et al. 2020
- (候選分類)卡瑞古菌屬(Kariarchaeum)Liu et al. 2020
- 嬌德古菌綱(Jordarchaeia)Sun et al. 2021
- 嬌德古菌目(Jordarchaeales)Sun et al. 2021
- 嬌德古菌科(Jordarchaeaceae)Sun et al. 2021
- (候選分類)嬌德古菌屬(Jordarchaeum)Sun et al. 2021
- 奧丁古菌綱(Odinarchaeia)
- 奧丁古菌目(Odinarchaeales)
- 奧丁古菌科(Odinarchaeaceae)
- (候選分類)奧丁古菌屬(Odinarchaeum)Tamarit et al. 2021
- 巴德爾古菌綱(Baldrarchaeia)Liu et al. 2020
- 巴德爾古菌目(Baldrarchaeales)Liu et al. 2020
- 巴德爾古菌科(Baldrarchaeaceae)Liu et al. 2020
- (候選分類)巴德爾古菌屬(Baldrarchaeum)Liu et al. 2020
- 索爾古菌綱(Thorarchaeia)
- 索爾古菌目(Thorarchaeales)
- 索爾古菌科(Thorarchaeaceae)(MBG-B)
- 赫爾莫德古菌綱(Hermodarchaeia)Liu et al. 2020
- 赫爾莫德古菌目(Hermodarchaeales)Liu et al. 2020
- 赫爾莫德古菌科(Hermodarchaeaceae)Liu et al. 2020
- (候選分類)赫爾莫德古菌屬(Hermodarchaeum)Liu et al. 2020
- 洛基古菌綱(Lokiarchaeia)corrig. Spang et al. 2015
- 赫爾古菌目(Helarchaeales)
- 洛基古菌目(Lokiarchaeales)Spang et al. 2015
- 洛基古菌科(Lokiarchaeaceae)Vanwonterghem et al. 2016
- (候選分類)洛基古菌屬(Lokiarchaeum)corrig. Spang et al. 2015 (MBGB, DSAG)
- MK-D1古菌科(MK-D1)
- (候選分類)普羅米修斯古菌屬(Promethearchaeum)corrig. Imachi, Nobu & Takai 2020
一些與阿斯加德古菌相關的、科層級的病毒已藉由宏基因組發現;[32][33][34]運用CRISPR片段比對病毒相對應的相鄰間隔原基序,這些病毒被認為與洛基古菌、索爾古菌、奧丁古菌、海姆達爾古菌有關。其中兩類病毒與感染細菌和細菌的、具有二十面體衣殼和螺旋尾的有尾噬菌體目相關;[32][34]另外兩類病毒則與具有二十面體衣殼但無尾的、感染細菌和細菌的多變去氧核糖核酸病毒域是遠親;[32][33]而第三類病毒則與帶有檸檬形病毒顆粒、專門感染古菌的病毒相關。[32][33]這些病毒在深海沉積物以及[32][34]黃石公園的地上熱泉中發現。[33]所有的這些病毒其序列與已知病毒的序列間的相似性非常低,但和其他已知的原核細菌相關,[35]而與真核生物的細菌無關。[36][32]
除了病毒外,現在透過CRISPR片段比對也已發現一些隱秘的、可能和洛基古菌門、索爾古菌門以及海姆達爾古菌門相關的可動遺傳因子,[32][37]這些可動遺傳因子並不轉譯成可辨識的病毒蛋白,且可能代表新種的病毒或質體。
張思豫; 盧中一; 黃文聰; 劉楊; 李猛. 古菌ESCRT系统研究进展. 微生物學報. 2020, 60 (6): 1304–1317.
張新旭; 李猛. 海洋沉积物中几类常见古菌类群的分布与代谢特征. 微生物學報. 2020, 60 (9): 1907–1921.
馬巧麗; 杜歡; 劉楊; 李猛. 红树林湿地硫酸盐还原菌的多样性及其参与驱动的元素耦合机制. 微生物學報. 2022, 62 (12): 4606–4627.
Taxon History. Genome Taxonomy Database. [6 December 2021]. (原始內容存檔於2021-11-01). Medvedeva S, Sun J, Yutin N, Koonin EV, Nunoura T, Rinke C, Krupovic M. Three families of Asgard archaeal viruses identified in metagenome-assembled genomes. Nature Microbiology. July 2022, 7 (7): 962–973. PMID 35760839. doi:10.1038/s41564-022-01144-6. Rambo IM, Langwig MV, Leão P, De Anda V, Baker BJ. Genomes of six viruses that infect Asgard archaea from deep-sea sediments. Nature Microbiology. July 2022, 7 (7): 953–961. PMID 35760837. doi:10.1038/s41564-022-01150-8.


