Gen liên kết
From Wikipedia, the free encyclopedia

Remove ads
Gen liên kết (linkage genes) là các gen cùng ở một nhiễm sắc thể, được di truyền cùng nhau cho thế hệ sau (xem hình bên).[1], [2]

Điều này có nghĩa là:
- Các tính trạng được quy định bởi các gen liên kết (linked genes) sẽ cùng biểu hiện với nhau ở một cơ thể, gây ra hiện tượng di truyền liên kết (genetic linkage);
- Các gen ở các nhiễm sắc thể khác nhau sẽ không cùng di truyền nên không cùng biểu hiện, là nguyên nhân của di truyền độc lập (independent genetics).[2]
Vì vị trí của một gen trên nhiễm sắc thể gọi là lô-cut (locus), nên có thể nói: các gen liên kết có locus ở cùng một nhiễm sắc thể, nguyên nhân gây ra hiện tượng di truyền liên kết.
Remove ads
Lược sử nghiên cứu
Thí nghiệm của Bateson, Saunders và Punnett (1905)
Hiện tượng gen liên kết được phát hiện và công bố đầu tiên ở Anh năm 1905,[3] nhờ William Bateson, Edith Rebecca Saunders - mẹ đẻ của Di truyền học thực vật Anh[4] và Reginald Punnett. Các nhà khoa học này nghiên cứu sự di truyền của hai gen sau đây ở cây đậu thơm (Lathyrus odoratus):
- Gen quy định màu hoa có alen trội P quy định màu tím, còn alen lặn p quy định màu đỏ;
- Gen quy định hình dạng của hạt phấn có alen trội L quy định hạt phấn dài, còn alen lặn l quy định hạt phấn tròn.
Khi thí nghiệm "lai theo kiểu Mendel" về hai tính trạng thuần chủng giữa dòng đậu thơm hoa tím, hạt phấn dài với dòng thuần hoa đỏ, hạt phấn tròn thì thu được 100 % cây F1 hoa tím, hạt phấn dài. Cho F1 tự thụ phấn với dự đoán sẽ được F2 phân li là 9 tím, dài: 3 tím, tròn: 3 đỏ, dài: 1 đỏ, tròn theo định luật Mendel mới được phát hiện lại cách đấy không lâu (năm 1900), thì kết quả thu được lại là 284: 21: 21: 55 (xem sơ đồ tóm tắt ở hình 2). Kiểm tra chỉ số χ2 (Khi bình phương) thì thấy rõ ràng các gen này không thể di truyền độc lập như Mendel đã chỉ ra. Do đó, họ khẳng định các gen này cặp đôi (coupling) hoặc kết nối (connection) nhau, rồi đưa ra giả thuyết sự lặp lại (reduplication) của các gen để giải thích.[5]

Thí nghiệm của Morgan (1910)
Thí nghiệm về gen liên kết giới tính
Sau đó ít lâu ở Mỹ, Morgan (Thomas Hunt Morgan) đã tình cờ phát hiện ra trong một quần thể ruồi giấm (Drosophila melanogaster) nuôi ở "phòng thí nghiệm ruồi" của mình có cá thể đực mắt trắng, khác hẳn tất cả các cá thể ruồi đã biết luôn có mắt đỏ. Ông thí nghiệm "lai theo kiểu Mendel" về 1 tính trạng thuần chủng: cho lai ruồi đực mắt trắng mới phát hiện này với ruồi cái mắt đỏ thuần chủng (P), thì thu được tất cả ruồi F1 mắt đỏ. Khi cho F1 giao phối với nhau đã thu được F2 có trung bình 75% số ruồi mắt đỏ: 25% số ruồi mắt trắng, nghiệm đúng tỉ lệ 3 trội: 1 lặn mà Mendel đã chỉ ra; nhưng có điều đặc biệt là ruồi con mắt trắng đều là giống đực (thí nghiệm 1, hình 3).[7], [8]
Bởi vậy, ông cho rằng màu mắt ruồi di truyền bị giới hạn bởi giới tính (sex limited inheritance).[7] Để kiểm tra điều này, ông cho lai ruồi đực mắt trắng với ruồi cái F1 (mắt đỏ dị hợp) thì thu được F2 gồm: 29 cái mắt đỏ: 132 đực mắt đỏ: 88 cái mắt trắng: 86 đực mắt trắng (thí nghiệm 2, hình 4),[7] khác hẳn định luật Mendel. Từ đó, dựa vào giả thuyết di truyền nhiễm sắc thể công bố năm 1903 của Theodor Boveri và Walter Sutton, ông phản đối giả thuyết "reduplication" (gen lặp lại) nói trên, mà Morgan coi là "thuần túy cơ học" (simple mechanical). Ông còn giả thuyết rằng: đặc điểm màu mắt được di truyền gắn với giới tính do gen này ở trên nhiễm sắc thể giới tính X và đưa ra ý tưởng về di truyền liên kết (genetic linkage). Trong giả thuyết này của ông (công bố năm 1911), nhiều gen (2 trở lên) cùng ở một nhiễm sắc thể thì chúng không di truyền độc lập nữa, mà di truyền liên kết với nhau. Ông cho rằng gen là có thật và "ánh xạ" từ vị trí xác định trên nhiễm sắc thể,[9] gọi là lô-cut (locus). Từ đó, ông hoàn thiện Học thuyết di truyền nhiễm sắc thể (giải Nobel 1934).[8]
Ngày nay, ta đã biết rằng: ở ruồi giấm thì alen quy định màu mắt đỏ là trội (Morgan coi đây là alen hoang dã trong tự nhiên và ông ký hiệu là dấu "+") định vị tại vùng không tương đồng của nhiễm sắc thể giới tính X, còn alen tương ứng quy định màu mắt trắng là đột biến lặn (ông kí hiệu là w, lấy chữ cái đầu của từ "white" là màu trắng). Nhiễm sắc thể giới tính Y của ruồi giấm không có lôcut này, nên sơ đồ thí nghiệm 1 tóm tắt là:
P: ♂ mắt trắng (XwY) × ♀ mắt đỏ (X+X+)
F1: ♀ X+Xw (đỏ) và ♂ X+Y (đỏ)
F2: 1 ♀ X+X+ (đỏ): 1 ♀ X+Xw (đỏ): 1 ♂ X+Y (đỏ): 1 ♂ XwY (trắng).
Điều này giải thích được tại sao thế hệ F2 phân li 3 trội: 1 lặn như ở định luật Mendel, nhưng cá thể nào mắt trắng thì phải là con đực bởi con đực chỉ cần 1 alen là màu mắt đã biểu hiện (hình 3).
Hiện tượng ở thí nghiệm 1 và 2 nói trên là trường hợp di truyền liên kết giới tính (sex linked).
Thí nghiệm về gen liên kết trên nhiễm sắc thể thường (1911)
Trong một thí nghiệm khác, Morgan đã xác định ở ruồi giấm thì tính trạng thân màu xám là trội (quy định bởi alen B) so với thân đen (quy định bởi alen b, black), còn cánh dài bình thường là trội (quy định bởi alen V) so với cánh cụt (quy định bởi alen v, vestigial). Sau khi tạo ra ruồi giấm đực dị hợp thân xám, cánh dài (BbVv), ông đã cho nó lai với ruồi cái thân đen, cánh cụt (bbvv) thì được đời con chỉ có 2 kiểu hình với tỉ lệ 1 xám, dài: 1 đen, cụt, không hề thấy 2 kiểu hình xám, cụt và đen, dài. Vì ruồi cái chỉ cho một loại trứng duy nhất là bv, nên ông khẳng định ruồi đực (BbVv) chỉ phát sinh hai loại tinh trùng là BV và bv, mà không có tinh trùng Bv và bV. Từ đó ông cho rằng các lôcut B/b và V/v cùng ở một nhiễm sắc thể nên không thể di truyền tách nhau như định luật Mendel. Thí nghiệm này chứng minh về gen liên kết rõ ràng nhất vào thời gian đó.
Sau này, đã xác định được vị trí của các lôcut mà Morgan gọi là B+/b và V+/v là ở nhiễm sắc thể số 2 của ruồi giấm (xem minh hoạ ở hình 5).
Để phân biệt với trường hợp các gen phân li độc lập, nhiều giáo trình Sinh học trên thế giới cũng như ở Việt Nam quy ước dùng kí hiệu các gen liên kết viết trên cùng một vạch (vạch này tượng trưng cho nhiễm sắc thể mang chúng). Ví dụ: ở giao tử (n) mà có gen A với gen B liên kết với nhau thì viết là AB. Ở tế bào soma (2n) có cặp nhiễm sắc thể tương đồng, nên nếu tinh trùng (giao tử đực) là AB còn trứng (giao tử cái) là ab, thì kiểu gen của hợp tử (2n) viết là AB//ab, hoặc AB/ab nhưng chuẩn nhất là kí hiệu như phân số có hai gạch ở hình 6.
Bản chất gen liên kết
Giờ đây, sau các sự kiện trên hơn 1 thế kỉ, nhờ sự phát triển của khoa học và công nghệ, người ta biết rõ rằng mỗi nhiễm sắc thể chứa một phân tử DNA mang nhiều gen. Các gen ở cùng phân tử DNA tạo nên một nhóm gen liên kết, số nhóm gen liên kết của mỗi loài bằng bộ nhiễm sắc thể đơn bội (n). Khi nhiễm sắc thể được truyền cho thế hệ sau, thì các gen cùng nhóm liên kết không phân li độc lập với nhau nữa.
Sau khi mô hình "chuỗi xoắn kép" DNA được công bố bởi Cơ-ric (Francis Harry Compton Crick) và Oat-sơn (James Dewey Watson), các nghiên cứu về nhiễm sắc thể dần dẫn đến hiểu biết về siêu cấu trúc của bào quan này ngày càng nhiều như mô hình của J. Herbert Taylor (1957), của Ris và Chandler (1963), của Kaufmann, McDonald, Whitehouse và Hastings (1965), mô hình sợi cuộn xoắn (folded fibre model) của Du Praw (1965), phát hiện về nuclêôxôm của A. Olins và C.L.F. Woodcock (1970) v.v.[10] Nhờ đó ta hình dung được siêu cấu trúc của nhiễm sắc thể - bào quan lưu giữ và bảo quản thông tin di truyền như bây giờ: tất cả các gen cùng thuộc về một phân tử DNA đã được cuộn xoắn phức tạp nhưng có tổ chức, lại được mê-tyl hóa thì sẽ cùng nhau truyền cho thế hệ sau (hình 7).
 Hình 3: Sơ đồ lai thuận (phép lai I) của Morgan.
Hình 3: Sơ đồ lai thuận (phép lai I) của Morgan. Hình 4: So sánh lai thuận (bên trái) và lai nghịch (phải) ở thí nghiệm của Morgan.
Hình 4: So sánh lai thuận (bên trái) và lai nghịch (phải) ở thí nghiệm của Morgan.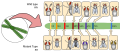 Hình 5: Phân bố một số lô-cut ở NST số 2 của ruồi giấm.
Hình 5: Phân bố một số lô-cut ở NST số 2 của ruồi giấm.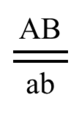 Hình 6: Kí hiệu chuẩn cho kiểu gen có 2 gen: A liên kết với B, và a liên kết với b.
Hình 6: Kí hiệu chuẩn cho kiểu gen có 2 gen: A liên kết với B, và a liên kết với b. Hình 7: Mỗi NST gồm khoảng 10 triệu đến 10 tỉ cặp base nitơ (bp) cuộn xoắn lại. Trong đó, mỗi gen khoảng 1000 đến 1 triệu bp.
Hình 7: Mỗi NST gồm khoảng 10 triệu đến 10 tỉ cặp base nitơ (bp) cuộn xoắn lại. Trong đó, mỗi gen khoảng 1000 đến 1 triệu bp.





Remove ads
Các kiểu gen liên kết
Qua trên, ta đã biết rằng các tính trạng di truyền liên kết với nhau là do các gen quy định những tính trạng đó ở trên cùng một nhiễm sắc thể. Tuy nhiên sự liên kết giữa chúng có thể theo nhiều kiểu đã phát hiện, đưa đến các kết quả khác nhau:
- Liên kết hoàn toàn (complete linkage) hay liên kết không hoàn toàn (incomplete linkage);
- Liên kết với nhiễm sắc thể thường (autosomal linkage) hay với giới tính (sex linkage).
- Các gen được xem là liên kết hoàn toàn khi chúng nằm sát nhau trên một phân tử DNA của nhiễm sắc thể và không có trao đổi chéo gây hoán vị các alen đang xét. Các gen được xem là liên kết không hoàn toàn khi chúng nằm xa nhau trên một phân tử DNA của nhiễm sắc thể và có xảy ra hoán vị gen tạo ra tái tổ hợp di truyền.[11]
- Các gen quy định những tính trạng thường (tính trạng không thuộc về giới đực, cái) mà lại ở nhiễm sắc thể giới tính (X hay Y, W hay Z) thì gây ra hiện tượng di truyền liên kết giới tính (sex linked). Nếu lô-cut đó ở nhiễm sắc thể X, người ta gọi là gen liên kết X (X-linked); nếu lô-cut đó ở nhiễm sắc thể Y, người ta gọi là gen liên kết Y (Y-linked). Ở người, gen liên kết X đã được phát hiện từ lâu, như gen đột biến lặn gây chứng máu khó đông (hemophilia), gen đột biến trội (gen MECP2) gây hội chứng Rêt do Andreas Rett (1924-1997) phát hiện. Khác hẳn với liên kết X, các tính trạng hình thái ngoài có liên kết Y hiện chưa được xác minh về cơ sở di truyền phân tử ở người. Một số tài liệu giáo khoa đã từng đưa ví dụ tính trạng hình thái ngoài được xem là liên kết Y như chứng hói đầu hay "túm lông tai" (hairy ears) chỉ ở đàn ông mới có, tuy nhiên vài năm gần đây đã được kiểm chứng lại.
- Chứng hói đầu đã từng được cho rằng do gen ở Y gây ra có liên quan tới cả sự phát triển của tóc và sự hình thành một số hormone. Ngày nay, người ta gọi gen này SOX21, còn có bí danh (alias) là SRY-Box21, nằm trên nhiễm sắc thể số 13, chứ không ở Y.[12]
- Từ lâu, đặc điểm có một túm lông ở tai đã được coi là tính trạng liên kết Y. Một số giáo trình di truyền học nước ngoài đã trình bày hiện tượng này như là một ví dụ điển hình về liên kết Y và gen "túm lông tai" gọi là gen holandric. Tuy nhiên, có nhà khoa học (Lee và cộng sự) cho rằng đây là một ngộ nhận, vì các nghiên cứu về giải trình tự nhiễm sắc thể Y hiện chưa định vị được gen này, hơn nữa đặc điểm này không phổ biến trên thế giới, mà chỉ gặp ở một số quần thể người ở Ấn Độ. Gần đây các nghiên cứu ở cấp độ phân tử cho rằng kiểu hình này là đột biến từ nhiều nhóm quần thể và có tính địa phương, nghĩa là các quần thể người Ấn Độ này có "ông tổ Adam" riêng.[13]
Tóm lại, gen liên kết là các gen cùng thuộc một phân tử DNA của một nhiễm sắc thể, các gen này có thể liên kết hoàn toàn hoặc liên kết không hoàn toàn, có thể liên kết giới tính hoặc không.
Remove ads
Nguồn trích dẫn
Wikiwand - on
Seamless Wikipedia browsing. On steroids.
Remove ads