Гаплогруппа R1a (Y-ДНК)
Y-хромосомная гаплогруппа Из Википедии, свободной энциклопедии

Гаплогруппа R1a (М420) — Y-хромосомная гаплогруппа, распространённая в Восточной и Центральной Европе, Средней и Южной Азии и Южной Сибири.
Эту страницу предлагается переименовать в «Гаплогруппа R1a». |
Этот грубый перевод статьи с неуказанного языка требуется улучшить (см. Рекомендации по переводу). Статья, целиком являющаяся машинным переводом, может быть удалена на основании критерия быстрого удаления С2. |
| Гаплогруппа R1a | |
|---|---|
 | |
| Тип | Y-ДНК |
| Время появления | 22 000 - 25 000 лет назад |
| Место появления | Южная Сибирь[1] |
| Предковая группа | R1 |
| Сестринские группы | R1b |
| Субклады | R1a1 (Z282 (Европа) и Z93 (Азия)), R1a2 |
| Мутации-маркеры |
R1a — L62, L63, L120, M420, M449, M511, M513 R1a1a — M17, M198, M512, M514, M515, L168, L449, L457, L566 |
R1a возникла около 22 тысяч лет назад[2] (по другим данным — около 25 тыс. лет назад[3]), предположительно в Азии (исходя из распространения линий R2 и R*). R1a-М420 делится на ветви R1a1-M459 и R1a2-YP4141.
Происхождение R1a1a1 (M417), древние миграции
Суммиров вкратце
Перспектива
R1a1a1-M417 это корневая и первая ветвь, пошедшая от гаплогруппы R1a-M420. Она разделилась между 25 и 22 тысячами лет назад. Это период последнего ледникового максимума. В исследовании 126 современных популяций со всей Евразии от 2014 года, проведённом Питером А. Андерхиллом и соавторами с участием 16 244 человек, авторы пришли к предположению о том, что «начальные эпизоды диверсификации гаплогруппы R1a, вероятно, произошли в окрестностях современного Ирана». Согласно расчётам Андерхилла и др. (2014), субклад R1a1a1-M417, диверсифицировался на линии, ведущие к субкладам R1a1a1b1a-Z282 и R1a1a1b2-Z93, около 5800 лет назад[3][4]. Раннеэнеолитический образец NEO113 из региона Среднего Дона (Голубая Криница) возрастом 7300 л. н. помещается в базальный субклад гаплогруппы R1a вместе с ранними образцами, связанными с комплексом шнуровой керамики, poz81 (R1a1a1a-CTS4385>CTS4385*, 2880-2630 до н. э., Obłaczkowo, Poland) и RISE446 (R1a1a1a-CTS4385>L664>S3479, 2829-2465 до н. э., Bergrheinfeld, Bavaria, Germany), что делает его самым ранним наблюдением этой базальной линии, о котором сообщалось до сих пор[5].
R1a показывает сильную корреляцию с индоевропейскими языками Южной и Западной Азии, Центральной и Восточной Европы[6][7]. Наибольшее распространение она имеет в Восточной Европе, Западной и Южной Азии и Центральной Азии. В Европе преобладает R1a1a1b1a-Z282, в то время как в Азии доминирует R1a1a1b2-Z93. Связь между Y-хромосомной гаплогруппой R1a1a-M17 и распространением индоевропейских языков впервые была отмечена Татьяной Зерджаль и её коллегами в 1999 году[8].
Взрывообразный рост числа потомков основателя субклады R1a1a1b2-Z93 произошёл в Южной Азии примерно 4,5—4 тыс. л. н. Расширение маркеров R1a1a1b2-Z93 в промежуток между 4500 и 4000 лет назад на несколько столетий предшествует угасанию и краху Индской цивилизации[9].
Согласно исследованию древней ДНК В. Нарасимхана и др. (2018), степные скотоводы являются вероятным источником Y-хромосомной гаплогруппы R1a в Индии[10]. К северу от ближневосточных регионов, уровни R1a1a начинают расти на Кавказе неравномерно. Несколько изученных популяций не показали никаких признаков R1a1a, в то время как самые высокие уровни, обнаруженные до сих пор в регионе, среди карачаевцев ок. 40 % и среди балкарцев ок. 35 % представлены доминирующей гаплогруппой R1a1a[7][11][12][13][14][15].
Теория происхождения из Азии
Т. Кивисилд в 2003 году предположил либо Южную, либо Западную Азию[16], а Мирабал в 2009 году указывал на Южную и Центральную Азии[6]. Другие исследователи предполагают происхождение R1a на территории современной Украины[17][18], в Средней Азии[19] и в Западной Азии[20][21].
В 2000 году предположено происхождение на территории Украины и постледниковое распространение R1a1 во время последнего ледникового максимума. Распространение генома по этой теории обеспечивалось в том числе благодаря экспансии курганной культуры в Европе и на Восток[22].
Спенсер Уэлс[англ.] предлагает центрально-азиатское происхождение, предполагая, что распределение и возраст R1a1 указывают на древнюю миграцию, соответствующую распространению Культуры курганных погребений в их экспансии из Евразийской степи[23].
В 2012 году предположено, что R1a1a выделилась в евразийских степях или на Ближнем Востоке и на Кавказе. Внутренняя и Центральная Азия, является перекрывающейся зоной для линий R1a1-Z280 и R1a1-Z93, что подразумевает раннюю зону дифференциации R1a1a-M198, предположительно произошла в пределах Евразийских степей или на Ближнем Востоке и на Кавказе, поскольку они лежат между Южной Азией и Центральной и Восточной Европой[24].
Три генетических исследования, проведённые к 2015 году, подтвердили курганную гипотезу Гимбутас. Согласно этим исследованиям, гаплогруппы R1b и R1a, теперь наиболее распространенные в Европе, были распространены из российских степей наряду с индоевропейскими языками. Исследования также обнаружили аутосомный компонент, присутствующий в современных европейцах, которого не было у неолитических европейцев, которые были бы введены с отцовскими линиями R1b и R1a, а также с индоевропейскими языками[25][26][27].
_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg)

 Европейский средний неолитический период. Культура ямочно-гребенчатой керамики, между 4200 г. до н. э. — ок. 2000 г. до нашей эры.
Европейский средний неолитический период. Культура ямочно-гребенчатой керамики, между 4200 г. до н. э. — ок. 2000 г. до нашей эры. Культура боевых топоров, с 2900 до н.э. – по 2350 до н.э.
Культура боевых топоров, с 2900 до н.э. – по 2350 до н.э. Трипольская культура — границы 1.
Трипольская культура — границы 1.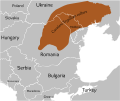 Трипольская культура — границы 2.
Трипольская культура — границы 2.




Связь с ямной культурой
Дэвид Энтони считал людей ямной культуры предками индоевропейцев[28][29]. Согласно исследованию 2015 года, массовая миграция людей ямной культуры имела место около 2500 лет до нашей эры. Её связывают с тем, что данная культура одной из первых массово научилась использовать лошадей для передвижения людей и перевозки грузов, а также активно применять бронзовое оружие. Генетики из Стэнфордского университета и др. (Poznik et al. 2016) выявили, что с демографической экспансией в Западную Европу в бронзовом веке связана не гаплогруппа R1a, а гаплогруппа R1b1a1a2a1a-L151/L11 (L23 и её субклады), в то время как субклад R1b-Z2103, представленный в ямных погребениях, в этой экспансии не участвовал[30][31].
Ямную культуру сменила катакомбная культура, у представителей которой также определили Y-хромосомную гаплогруппу R1b, а не R1a[32][33].
Исследователи отмечают, что R1a и R1b могли распространиться в Европу с Востока после 3000 лет до нашей эры[29]. Однако при исследованиях все семь образцов из ямной культуры принадлежали к R1b1a1a2-M269, а R1a1a не была обнаружена в образцах, отнесённых к ямной культуре[29]. Это поставило вопрос — откуда возникла R1a1a в культуре боевых топоров, учитывая что ямная культура не являлась носителем данной субклады.
Семёнов и Булат спорят о происхождении R1a1a в культуре боевых топоров, отмечая, что несколько исследований указывают на присутствие R1a1a в генетике данной культуры[34][35][36][37]. Некоторые исследователи полагают, что R1a попала на Балканы через Анатолию, а уже оттуда распространялась дальше на северо-запад, до культуры рёссен, затем на восток от трипольской культуры до ямной культуры и афанасьевской культуры. При этом исследователи признают — R1a отсутствует в промежуточных культурах между Ближним Востоком, Анатолией и Балканами. Аско Парпола, исследуя эту проблему, предположил, что трипольская культура соотносится с поздним вариантом праиндоевропейского этноса. Он отмечает, что в трипольской культуре, возможно, изобретено колесо, а также что, вероятно, люди трипольской культуры была захвачены или ассимилированы носителями праиндоевропейского языка в 4-м тысячелетии до н. э., после чего культура распространилась до Понтийской степи к 3400 годам до нашей эры и в результате развития перетекла в ямную культуре[38].

Закавказье и западно-европейское происхождение, возможное влияние Индской цивилизации
Часть генетической составляющей в жителях Южной Азии происходит от западно-европейского населения, и некоторые исследователи предполагали, что субклад R1a1a1b2-Z93, возможно, прибыл в Индию через Иран[39] и расширился там во время Хараппской цивилизации[40][41].
В 2015 году предположено[42], что корни R1a1a1b2-Z93 лежат в Западной Азии и её распространение вместе с L342.2 проходило в Юго-Восточном направлении от Закавказья до Южной Азии"[39], мотивируя это археологическими данными о восточной популяции Западной Азии в 4-м тысячелетии до нашей эры, кульминацией которого стали так называемые миграции куро-араксской культуры в период после Урука IV[англ.] (период истории Месопотамии между 4000 — 3100 лет до нашей эры)[39]. Тем не менее Лазаридис отметил, что в куро-аракский образец I1635 была привнесена Y-хромосомная гаплогруппа R1b1-M415 (xM269)[43] также называемая R1b1a1b-CTS3187[44].
Согласно результатам исследования 2014—2015 годов[45], диверсификация-отделение R1a1a1b2-Z93 и процессы урбанизации в долине Инда произошли 5600 лет назад, и географическое распределение R1a1a1b2a1~-M780 (нижестоящего к R1a1a1b2a-Z94)[46] может отражать это.
В 2016 году отмечено[47], что «произошло поразительное расширение» субклады R1a1a1b2-Z93 в промежуток между 4500—4000 л. н., что на несколько столетий предшествует угасанию, краху Индской цивилизации[41].

Теория южно-азиатского происхождения
В исследовании 2009 года указывается в качестве возможной «прародины» гаплогруппы как Южная, так и Центральная Азия[48].
Южноазиатские популяции имеют наивысшее разброс микросателлитов в пределах R1a1a[48][48][49][50][51], и последующих, более поздних датировок на ближайшего общего предка. А R1a1a присутствует как среди высших каст (брахманов), так и низших каст, хотя присутствие значительно выше среди брахманов[51]. Исходя из этих данных, некоторые исследователи пришли к выводу, что R1a1a возникла в Южной Азии[52], исключая существенный генетический приток от индоевропейских мигрантов[52][53][54].
Однако это разнообразие и последующие более ранние датировки на выявление ближайшего общего предка также могут быть объяснены исторически большим числом населения, что увеличивает вероятность диверсификации и изменения микросателлитов[55][56][57]. R1a1 и R2, возможно, распространялись в Южную Индию из юго-западного региона Азии несколько раз[53].
В 2017 году отмечено[58], что R1a в Южной Азии наиболее вероятно распространилась от одного центрального пучка источников в Центральной Азии, по крайней мере — трёх. И, вероятно, эти генетические линии в дальнейшем развивались в границах Индийского субконтинента, что согласуется с несколькими волнами миграций[55].

Топология
Топология R1a выглядит следующим образом[59][60][61][62].
Согласно работе Tatiana Karafet et al. (2014), быстрый процесс диверсификации гаплогруппы K-M526, вероятно, произошел в Юго-Восточной Азии с последующими расширениями на западе предков гаплогрупп R и Q[62][63].
- P P295/PF5866/S8 (ранее известная как K2b2).
- R (R-M207)[64][65]
- R*
- R1 (R-M173)
- R1*[65]
- R1a (M420)[65] (Eastern Europe, Asia)[66]
- R1a*[64]
- R1a1[65] (M459/PF6235,[65] SRY1532.2/SRY10831.2[65])
- R1a1 (M459)[64][65]
- R1a1b (YP1272) https://web.archive.org/web/20150428004519/http://forum.molgen.org/index.php?topic=7835.0
- R1a1a (M17, M198)[65]
- R1a1a1 (M417, page7)[65]
- R1a1a1a (CTS7083/L664/S298)[65]
- R1a1a1b (S224/Z645, S441/Z647)[65]
- R1a1a1b1 (PF6217/S339/Z283)[65]
- R1a1a1b2 (F992/S202/Z93)[65] [R1a1a2*] (Z93, M746)(Asia)[67]
- [R1a1a1c] (M64.2, M87, M204)[68]
- [R1a1a1d] (P98)[68]
- [R1a1a1d2a][70]
- [R1a1a1e] (PK5)[68]
- R1a1a1 (M417, page7)[65]
- R1b (M343) (Western Europe)
- R2
Палеогенетика
Суммиров вкратце
Перспектива
Самые древние известные R1a обнаружены у следующих двух (возраст около 10800 лет[72]):
(субклад R1a1b~-YP1272>YP1301) определена у представителя культуры Веретье из могильника Песчаница 1 PES001 (10785—10626 лет назад) на озере Лача (Каргопольский район Архангельской области)[73].
У мезолитчика I1819, жившего 10825—10561 лет назад Василевка-на-Днепре, определена Y-хромосомная гаплогруппа R1a-M420>R1a1-M459>M459*.[74]
Гаплогруппа R1a обнаружена в останках следующих культур и народов: культура боевых топоров[75][76][77], Культура полей погребальных урн[78], синташтинская культура[79], андроновская культура[80], пазырыкская культура[81], тагарская культура[80], таштыкская культура[80], у жителей древнего Танаиса (город)[82], в таримских мумиях[83], у аристократов Хунну[84], в двух древних останках хазар.
У представителей фатьяновской культуры определена Y-хромосомная гаплогруппа R1a1a1-M417, при этом у половины образцов удалось уточнить субклад R1a1a1b2-Z93[73].
Линия R1a1a1b2a1~-Y3, предковая для субклада R1a1a1b2a1a-L657, в настоящее время распространëнного особенно в Индии в высшей варне — у браминов, найдена у образца I6561 в Александрии у реки Оскол, в Харьковской области (Alexandria, Ukraine_MBA, Украина). Датировка захоронения 2134—1950 гг. до н. э.[85][86]
| Номер теста | Захоронение | Датировка | Субклады | Культура |
|---|---|---|---|---|
| uzoo74 | Южный Олений остров на Онежском озере | ок. 8 тыс. лет назад | R1a1-SRY10831.2 | мезолит [87] |
| — | Глазковский некрополь | 8000—7000 лет назад | R1a1-M17 | неолит[88] |
| nik002 | Никульцынский могильник[89] | 2865—2500 гг. до н. э. | R1a-Z645 | фатьяновская культура [90] |
| vor005 | Воронковский могильник[89] | 2840—2343 гг. до н. э. | R1a-M417 > R1a-Z645 | фатьяновская культура[90] |
| tim008 | Тимофеевский могильник[91] | 2832—2473 гг. до н. э. | R1a-Z645 | фатьяновская культура[90] |
| vor003 | Воронковский могильник[89] | 2573—2466 гг. до н. э. | R1a-Z645 | фатьяновская культура[90] |
| ber1 | Бергсгравен (Bergsgraven), Швеция | — |
R1a1a1b1-Z283 |
культура боевых топоров[92] |
| kzb005 | Казбурун 1 | 1880—1690 гг. до н. э. | R1a-M459, R1a-Z283, Z280 | срубная и алакульская[93] |
| kzb008 | Казбурун 1 | 1880—1690 гг. до н. э. | R1a-M459, R1a-Z283, Z280, S24902? | срубная и алакульская[93][94] |
| kzb002 | Казбурун 1 | 1875—1665 гг. до н. э. | R1a-M459, R1a-Z93, YP5585 | срубная и алакульская[93] |
| kzb003 | Казбурун 1 | 1765—1630 гг. до н. э. | R1a-M459, R1a-Z93, Z94 | срубная и алакульская[93] |
| mur002 | Мурадым 8 | — | R1a-M459 | срубная и алакульская[93] |
| mur003 | Мурадым 8 | 1880—1685 гг. до н. э. | R1a-M459, R1a-Z93 | срубная и алакульская[93] |
| kzb007 | Казбурун 1 | 1755—1630 гг. до н. э. | R1a-M459, R1a-Z93, Z94, Z2124, Z2125, Z2123, Y934, BY30762? | срубная и алакульская[93] |
| mj31 | Дикий Сад | 1281—1058 гг. до н. э. | R1a-Z645 | киммерийцы[95] |
| cim357 | Глиное Сад | 914—805 гг. до н. э. | R1a-M459, R1a-Z93 | киммерийцы[93] |
| mj32 | Бидыло | 764—430 гг. до н. э. | R1a2c-B111 | киммерийцы[95] |
| scy193* | Глиное | — | R1a-M459, R1a-Z93, Z94, Z2124, Z2125, Z2123, Y934, BY30762, BY30764? | скифы[93] |
| chy002 | Чёрный Яр | 65 — 220 гг. н. э. | R1a-M459, R1a-Z93, Z94, Z2124, R1a1a1b2a2-Z2122, Y57, Y52 | сарматы[93] |
| tem002 | Темясово | 125—240 гг. н. э. | R1a-M459, R1a-Z93, Z94, Z2124, Z2125, S23592, YP1558, Y73758, YP5844, FGC48758 | сарматы[93] |
| tem003 | Темясово | 130—320 гг. н. э. | R1a-M459, R1a-Z93, Z94, Z2124, Z2125, Z2123, YP3920 | сарматы[93] |
Скелеты отца и двух его сыновей с археологического объекта, обнаруженного в 2005 году недалеко от Ойлау (Саксония-Анхальт, Германия) и датируемых примерно 2600 годом до н. э., показали положительный результат для маркера Y-SNP SRY10831.2. Таким образом предковая клада присутствовала в Европе по меньшей мере 4600 лет назад и была связана с одним из археологических памятников культуры боевых топоров (шнуровой керамики)[96]. Номер Ysearch для останков Eulau — 2C46S. У индивидов из Ойлау[нем.] и у образцов из Волосово-Даниловского могильника (R1a-S224, фатьяновская культура, Ярославская область) значения аллелей по 10 локусам совпадают, а по 4 локусам различаются на единицу[97].
Также гаплогруппа R1a была выявлена у представителей межовской[98], хвалынской, срубной, потаповской, полтавкинской[99], тшинецкой[100]. археологических культур, на стоянке Афонтова гора (бронза)[98], у скифов[101][102][103], у населения срубной культурно-исторической общности[103], среди аланских захоронений (субклад Z2124 — R1a1a1b2a2)[104], у населения салтово-маяцкой культуры (субклад Z2124 — R1a1a1b2a2)[104], у одного представителя культуры шнуровой керамики (боевых топоров) из Эсперштедта и у двух из саксонской пещеры Лихтенштейн, у одного представителя культуры полей погребальных урн из Хальберштадта и у двух из Эйлау[87][105]. Гаплогруппа R1a1 прослежена у обитателей верховий Западной Двины с 5120±120 л. н. (Сертея VIII) до VIII—V веков до н. э. (городище Анашкино)[106].
Европа
Гаплогруппа R1a наиболее часто встречается у водь, лужичан, чехов, словаков, поляков, русских, белорусов, украинцев, литовских татар[107][108], кубанских ногаев[109][110], крымских татар[111][112][113][114], карачаево-балкарцев[115] и некоторых других популяций. Причём для балтийских и славянских народов более характерна европейская ветка R1a-M458, а для тюркских — азиатская R1a-Z93[116], хотя и среди тюрков Европы достаточно распространены европейские ветки R1a[117].
Центральная Азия и Алтай
Суммиров вкратце
Перспектива
На Алтае, в Центральной и Южной Азии преобладает R1a-Z93[118].
У тюркского народа хотонов отмечена самая высокая доля носителей R1a1a в мире — 83 %[119], у киргизов R1а1а — 63,5 %[120]. Значительные доли R1a1a отмечены у тюркских народов азиатской части России и центральной Азии: алтайцев — до 58 %[121], шорцев — 56,9 %, телеутов — 53,3 %[122], теленгитов (более 50 %)[123].
В Афганистане R1a1a встречается у 51 % пуштунов и 45 % у таджиков[124], реже — среди хазарейцев (7 %). У хакасов — 28 %, у казахов родов — ошакты 26,7 %, суан — 31,7 % и кожа 35 %. У узбеков ~25 %[125][124], у уйгур — 31 %. У тувинцев — 12 % (наибольшая концентрация у северо-восточных — 26 %), у бурят — 4 %. В Таджикистане представлена по-разному: от 16 % у ягнобцев и 19 % у таджиков Душанбе до 68 % у ишкашимцев (N=25), что может объясняться дрейфом генов[124]:2, 5. Примечательно, что население современного Ирана, говорящее на одном из основных индоевропейских языков (фарси), по-видимому, испытало незначительное генетическое влияние со стороны популяций, несших R1a (среди иранцев Тегерана встречается в 4 % случаях, у иранцев Шираза и Исфахана не обнаружена)[124]:2, 5.
R1a-Z2125 наиболее часто встречается среди киргизов и афганских пуштунов (>40 %), у других народов Афганистана встречается с частотой >10 %. Парагруппа R1a-Z93* наиболее распространена (>30 %)[118]:126 среди коренных народов южносибирского Алтая (алтайцы, теленгиты, кумандинцы)[121], но также встречается в Кыргызстане (>6 %) и среди иранских народов (1—8 %)[118]:126.
Южная Азия
В Южной Азии R1a1a часто наблюдается с высокой частотой в ряде демографических групп.
В Индии высокие частоты этой гаплогруппы наблюдаются у западных бенгальских брахманов[англ.] (72 %) на восток, лоханов (60 %) на западе, кхатри[англ.] (67 %) на севере и айенгары[англ.] (31 %) на юге. Она также была найдена у нескольких южно-индийских дравидийских племён адиваси, включая ченчу[англ.] (26 %), валмики из Андхра-Прадеша и калларов[англ.] в Тамилнаде, что говорит о том, что R1a1a широко распространена у племенных южных индийцев.
Кроме того, исследования показывают высокие проценты в региональных группах, таких как Манипури (50 %), на крайнем северо-востока и среди пенджабцев (47 %) на крайнем северо-западе
В Пакистане он встречается у 71 % среди племени Моханны в провинции Синд на юге и 46 % среди балтийцев Гилгит-Балтистан на севере. Среди сингальцев Шри-Ланки 23 % были обнаружены как R1a1a (R-SRY1532). Индусы района Читван в регионе Тераи Непал показывают его на уровне 69 %[52][53][54][126].
Восточная Азия
В Восточной Сибири R1a1a встречается среди некоторых коренных этнических групп, включая камчатцев и чукчей, и достигает у ительменов 22 %[11].
Западная Азия
R1a1a был обнаружен в различных формах, в большинстве районов Западной Азии, в самых разных концентрациях. От почти не присутствующих в таких районах, как Иордания, до гораздо более высоких уровней в некоторых частях Кувейта, Турции и Ирана. Племена бедуинов Шаммар в Кувейте показывают самую высокую частоту на Ближнем Востоке — до 43 %[127].
Известные представители гаплогруппы R1a
- По данным генетиков у представителей династии Арпадов, Белы III и у Ласло I Святого[128], определена Y-хромосомная гаплогруппа R1a (Z93-Z2123>Y2632>SUR51 субклад[129]) и у Белы III митохондриальная гаплогруппа H1b[130].
Примечания
Литература
Ссылки
Wikiwand - on
Seamless Wikipedia browsing. On steroids.