Razvoj središnjeg živčanog sustava
Razvoj središnjeg živčanog sustava From Wikipedia, the free encyclopedia

Središnji živčani sustav (lat. systema nervosum centrale ili pars centralis systematis nervosi) razvija se iz ektoderma. Početkom 3. tjedna embrionalnog života javlja se neuralna ploča kao zadebljanje ektoderma s dorzalne (stražnje) strane zametnog štita. Potom se lateralni (postranični) rubovi neuralne ploče uzdignu u neuralne nabore koji se dalje uzdižu, primiču i konačno spajaju tvoreći neuralnu cijev. Kranijalni i kaudalni kraj te cijevi su prvo otvoreni gdje je prednji otvor neuroporus anterior (zatvara se 25. dan), a stražnji neuroporus posterior (zatvara se 27. dan). Stijenka neuralne cijevi izgrađena je od nediferenciranih stupićastih neuroepitelnih stanica koje su usmjerene od šupljine prema površini neuralne cijevi i raspoređene u jednom sloju tako da su njihove jezgre položene u više redova (tzv. pseudostratificirani stupićasti epitel). Od njih se umnažanjem (proliferacijom) razviju svi neuroni i makroglija (astrociti i oligodendrociti) središnjeg živčanog sustava čije su sastavnice:[1][2]
| Središnji živčani sustav |
Mozak | Prosencephalon | Telencephalon |
Rhinencephalon, Amygdala, Hippocampus, Neocortex, Bazalni gangliji, Lateralne moždane komore | |
| Diencephalon |
Epithalamus, Thalamus, Hypothalamus, Subthalamus, Hipofiza, Epifiza, Treća moždana komora | ||||
| Moždano deblo | Mesencephalon |
Tectum, Moždani krakovi, Pretectum, Vodovod srednjeg mozga (lat. aqueductus mesencephali) | |||
| Rhombencephalon | Metencephalon |
Most (lat. pons), Mali mozak (lat. cerebellum) | |||
| Myelencephalon | Produžena moždina | ||||
| Kralježnična moždina | |||||

Histogeneza

Histogeneza je proces uspostavljanja histološke (tkivne) građe središnjeg živčanog sustava. Histogenetski procesi uzrokuju tri temeljne promjene neuralne cijevi embrija: a) intenzivno se povećava cjelokupna razvojna osnova središnjeg živčanog sustava, b) promjene oblika različitim rastom dijelova cijevi pa se neuralna cijev na nekoliko mjesta pregiba i mijenja oblik, a to postupno nastajanje vanjskog oblika odraslog mozga je morfogeneza, c) promjenama građe stijenke neuralne cijevi nastaju privremene embrionalne ili fetalne zone.
Ključna promjena oblika neuralne cijevi je pojava moždanih mjehurića, što omogućuje temeljnu regionalnu podjelu mozga. Razlike intenziteta, trajanja i posebnih obilježja histogenetskih procesa u svakom pojedinom području moždane osnove omogućuju nam podjelu razvojnog procesa u niz razvojnih stadija, faza i razdoblja. Ta razvojna razdoblja potom služe kao vremenski pokazatelji za praćenje histogenetskih procesa i kritičnih razdoblja, vulnerabilnosti i stupnja razvojnog poremećaja nakon djelovanja različitih patogenetskih činitelja. Pored toga, navedene razlike intenziteta, trajanja i posebnih obilježja histogenetskih procesa dovode do oblikovanja posebnih slojeva stijenke embrionalnog i fetalnog mozga.Ti slojevi su embrionalne zone, što služe kao prostorni pokazatelji histogenetskih procesa. Tri embrionalne zone su univerzalne jer ih imaju svi dijelovi neuralne cijevi. Poredane od šupljine neuralne cijevi prema pijalnoj (vanjskoj) površini: ventrikularna zona (VZ), intermedijalna zona (IZ) i marginalna zona (MZ). U stijenci fetalnog telencefalona razviju se još tri posebne zone: subventrikularna zona (SVZ), subplate zona (SP) i kortikalna ploča (KP). MZ, KP i SP zajedno tvore razvojnu osnovu moždane kore.
Histogenetski procesi
Histogenetske procese u središnjem živčanom sustavu dijelimo u dvije temeljne skupine:
- Progresivni histogenetski procesi - od malog i jednostavnog prema većem i složenijem: a) proliferacija (umnožavanje stanica nizom uzastopnih mitoza), b) migracija (putovanje stanica od mjesta posljednje mitoze do ciljnog područja u kojem se konačno smjeste), c) diferencijacija morfološkog fenotipa neurona tj. izrastanje dendrita (dendrogeneza) i aksona (aksonogeneza) i dostizanje odraslog oblika i veličine some (tijela neurona), d) diferencijacija kemijskog fenotipa tj. razvoj neurotransmiterske specifičnosti neurona, e) razvoj sinapsi (sinaptogeneza) čime se uspostavljaju specifični neuronski sustavi i krugovi.
- Procesi razvojne reorganizacije -tijekom razvoja, strukturno-funkcionalno ustrojstvo mozga se nekoliko puta temeljito promjeni, a te promjene nazivamo procesima razvojne reorganizacije. Primjerice, embrionalne i fetalne zone su privremene tvorbe i nema ih u odraslom mozgu, a neke vrste neurona (npr. subplate neuroni i Cajal-Retziusove stanice) i glije (npr. radijalna glija) postoje samo u fetalnom mozgu, gdje imaju specifične razvojne funkcije. Stanice, aksoni i sinapse isprva se prekomjerno stvaraju, a potom velik dio njih nestane. Tipični reorganizacijski procesi su sljedeći: a) prirodna smrt prekobrojnih neurona, b) povlačenje i premještanje aksona ili aksonskih ogranaka (nestanak privremenih projekcijskih veza između neurona), c) smanjivanje broja prekobrojnih dendritičkih trnova, d) nestanak privremenih i smanjivanje broja prekobrojnih sinapsi, e) promjene neurotransmiterskog fenotipa neurona.
Razvoj mozga - moždani mjehurići


Nejednako odvijanje histogenetskih procesa dovodi do pojave tri uzastopna proširenja prednjeg kraja neuralne cijevi - to su primarni moždani mjehurići: prosencephalon (prednji mozak), mesencephalon (srednji mozak) i rhombencephalon (stražnji mozak). U 5. embrionalnom tjednu nastaju sekundarni moždani mjehurići jer se prosencephalon podijeli na telencephalon (krajnji mozak) i diencephalon (međumozak), a rhombencephalon se podijeli na metencephalon i myelencephalon. Od tih mjehurića se razviju glavna područja odraslog mozga:
- Telencephalon: od parnih telencefaličkih mjehurića razviju se moždane polutke (hemisfere) čiju površinu prekriva tanka moždana kora (cortex cerebri), a unutrašnjost čine bijela tvar velikog mozga (substantia alba cerebri), bazalni gangliji telencefalona (nucleus caudatus, putamen, claustrum i corpus amygdaloideum) i lateralne moždane komore. Od središnjeg, neparnog dijela (telencephalon impar) što spaja lijevi i desni telencefalički mjehurić, razviju se strukture mediobazalnog telencefalona i lamina terminalis, a u tom se području razviju i tri komisurna sustava telencefalona: corpus callosum, commissura anterior i commissura hippocampi.
- Diencephalon: tri privremena uzdužna žlijeba podijele diencefalon u četiri uzdužne zone od kojih se razviju epithalamus, thalamus, hypothalamus i subthalamus. Nadalje, i globus pallidus razvije se od diencefaličkog mjehurića. Od šupljine diencefaličkog mjehurića nastaje III. moždana komora.
- Mesencephalon: od ovog mjehurića, što ostaje nepodijeljen, ventralno se razviju pedunculi cerebri, a dorzalno ploča s četiri kvržice, lamina quadrigemina, što oblikuje tectum mesencephali (nju čine dva gornja i dva donja kolikula). Šupljina mjehurića toliko se suzi da nastane uski kanal, aquaeductus mesencephali (Sylvii) što spaja III. s IV. moždanom komorom.
- Isthmus rhombencephali: to je suženje neuralne cijevi što čini najrostralniji dio rombencefalona i označava prijelaz rombencefalona u mezencefalon; od njega se razviju gornji kraci malog mozga (pedunculi cerebellares superiores) i velum medullare superius.
- Metencephalon: to je rostralni dio rombencefalona od kojeg se razviju most (pons) na ventralnoj i mali mozak (cerebellum) na dorzalnoj strani.
- Myelencephalon: to je kaudalni dio prvotnog rombencefalona od kojeg se razvije produljena moždina (medulla oblongata). Od šupljina rombencefaličkog mjehurića kasnije nastaje IV. moždana komora. Od preostalog, kaudalnog dijela neuralne cijevi razvije se kralježnična moždina (medulla spinalis).
Razvitak površine mozga
Razvoj površine mozga tj. korteksa promatramo kroz filogenezu. Na bazalnoj površini nalazi se najstariji dio kore mozga, paleocortex. Na njemu se već vrlo rano uočava lagano zadebljanje koje se razvija u njušnu lukovicu, bulbus olfactorius. Iz središnjeg dijela svake hemisfere lateralno usmjerenim rastom nastaju parijetalni i temporalni režanj, od kojega se bazalni dijelovi (buduća area entorhinalis et hippocampus) razvijaju najranije i definiraju kao archicortex (ili archipallium, gr. archaios – star). Svi kasnije razvijeni dijelovi kore mozga ubrajaju se u neocortex (neopallium, gr. neos – nov, mlad). Neocortex ima šest jedinstvenih slojeva te se označuje i kao isocortex. Filogenetski stariji paleocortex i archicortex pokazuju heterogenu laminaciju i označuju se kao allocortex.
Daljnje povećanje kortikalne površine zbiva se preko nabiranja, nastanak vijuga lat. gyri (sulcificiranje) te uz njih priliježuće brazde, lat. sulci. Kao prva brazda nastaje sulcus lateralis cerebri između frontalnog i temporalnog režnja. Nakon toga pojavljuje se središnja brazda, sulcus centralis, kao granica između frontalnog i parijetalnog režnja. Na medijalnoj strani oblikuju se daljnje dvije brazde, sulcus pariteooccipitalis kao granica parijetalnoga i okcipitalnog režnja, te sulcus calcarinus. Na kraju prvog trimestra trudnoće mozak i kralježnična moždina su već u velikoj mjeri poprimili odrasli oblik. Jedino je upadljiv relativno neproporcionalno velik volumen ventrikularnog prostora. S rastom snopova aksona smanjuje se lumen na veličinu u odrasle osobe.
 Vijuga (gyrus) i žlijeb (sulcus).
Vijuga (gyrus) i žlijeb (sulcus). Vijuge i žljebovi mozga s vanjske (lateralne strane).
Vijuge i žljebovi mozga s vanjske (lateralne strane). Vijuge i žljebovi s unutrašnje (medijalne) strane.
Vijuge i žljebovi s unutrašnje (medijalne) strane.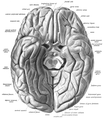 Vijuge i žljebovi s bazalne (donje) strane.
Vijuge i žljebovi s bazalne (donje) strane. Lateralni pogled.
Lateralni pogled. Različiti stadiji razvoja moždane površine.
Različiti stadiji razvoja moždane površine.






Postnatalni razvitak i težina mozga
Nakon završetka morfogeneze dolazi još tijekom trudnoće, dojenačkoga razdoblja i ranog djetinjstva do daljnjeg sazrijevanja mozga i kralježnične moždine. Međutim, nakon rođenja ne dolazi do daljnjeg povećanja broja neurona, već do oblikovanja tipične morfologije i stvaranja sinapsi. Rast dendrita, stvaranje dugih veza, mijelinizacija i povećanje ekstracelularnog prostora dovode mozak do njegove odrasle veličine. Težina mozga u odraslih je 1.3 do 1.4 kg (apsolutna težina mozga) ili 20 g po kg tjelesne težine (relativna težina mozga).
Proliferacija tijekom histogeneze
Proliferacija se odvija u ventrikularnoj i subventrikularnoj zoni. Proliferacija stanica odvija se u ventrikularnoj zoni (VZ), što je u ranim stadijima smještena uz šupljinu neuralne cijevi, a kasnije uz šupljinu fetalnih moždanih komora. Neuroepitelne stanice ventrikularne zone i srodne joj subventrikularne zone preteče su svih neurona i makroglije središnjeg živčanog sustava. Nakon intenzivne proliferacije u VZ, vanjski nastavci neuroepitelnih stanica stvaraju novu, površinsku marginalnu zonu (MZ). Tijekom daljnje intenzivne proliferacije, mnoge postmitozne stanice napuštaju VZ i putujući prema površini oblikuju novu, intermedijalnu zonu (IZ) na granici VZ i MZ. Prve stanice intermedijalne zone su nezreli, isključivo postmitotički neuroni, što se nikad više ne dijele.
Migracija

Svi se neuroni prođu kroz posljednju mitozu na jednom mjestu - u VZ ili SVZ uz ventrikularnu površinu, a potom ti nezreli postmitotički neuroni moraju otputovati do svojeg odredišta na drugom mjestu. Riječ je o procesu neuronske migracije. Neuroni migriraju kroz vrlo složenu tkivnu mrežu intermedijalne i (u telencefalonu iza 12. tjedna) subplate zone pa im pomažu kao vodiči radijalne glijalne stanice razapete poput užadi od ventrikularne do pijalne površine fetalnog mozga. Migrirajući neuroni zapravo pužu uz radijalnu gliju (slika), pa se stoga i migracija u pravilu odvija u radijalnom smjeru, prema površini neuralne stijenke. Stanice rođene u različito vrijeme, ali na istom mjestu u ventrikularnoj zoni telencefalona, migriraju duž istih radijalnih vodiča i završavaju u osnovi moždane kore jedne iznad drugih. Proliferacijska zona je mozaik proliferacijskih jedinica, a svi postmitotički neuroni nastali iz jedne takve jedinice su poliklon i migriraju duž istih radijalnih vodiča. Tako neuroni pronalaze pravi put kroz intermedijalnu i subplate zonu do svog konačnog odredišta u kortikalnoj ploči. Stoga svi postmitotički neuroni nastali u jednoj proliferacijskoj jedinici na kraju oblikuju morfološki prepoznatljiv stupićasti skup neurona u kortikalnoj ploči - ontogenetsku ili embrionalnu kolumnu. To znači da je vodoravni položaj neurona (pripadnost određenoj embrionalnoj kolumni) određen međusobnim rasporedom proliferacijskih jedinica u ventrikularnoj zoni.[3][4]
Razvoj sinapsi

Pojava i razvoj neuronskih veza ključni je histogenetski proces u središnjem živčanom sustavu. U tom procesu razlikujemo: a) rast presinaptičkih aksona, b) razvoj postsinaptičkog elementa i c) razvoj samih sinapsi tj. sinaptogenezu. Većina presinaptičkih aksona u fetalnom mozgu su aferentni aksoni iz udaljenih područja. Općenito, monoaminske skupine neurona moždanog debla (što sintetiziraju noradrenalin, dopamin ili serotonin) sazrijevaju vrlo rano i svoje projekcijske aksone šalju u druga područja mozga pa tako i u fetalni telencefalon. Slično tome, aksoni acetilkolinskih neurona bazalnog telencefalona i talamokortikalni aksoni rano urastaju u osnovu moždane kore. Ti aksoni mogu sadržavati neurotransmitere i prije no što uspostave prve sinapse, a glavna postsinaptička mjesta u ranom fetalnom mozgu su dendriti (rane sinapse su obično aksodendritičke). U fetalnom telencefalonu čovjeka, prve sinapse pojave se već krajem 8. tjedna trudnoće, kad većina budućih neurona moždane kore još nije niti rođena. Nadalje, te rane sinapse prvo se pojave u dvije odvojene zone: iznad kortikalne ploče (u marginalnoj zoni) i ispod nje (u subplate zoni), a tek u kasnijim stadijima i u samoj kortikalnoj ploči. No, valja naglasiti da su sve tri zone razvojna osnova odrasle moždane kore. Sinaptogeneza se u moždanoj kori ljudskog fetusa intenzivno odvija tijekom dva razdoblja: između 13. i 16. tjedna i između 22. i 26. tjedna trudnoće; vjerojatno postoji i treće, kasno razdoblje moćne sinaptogeneze, u doba početnog oblikovanja konačne slojevite građe moždane kore.
Nestajanje prekobrojnih sinapsi
Prekomjerno stvaranje sinaptičkih elemenata i samih sinapsi te njihovo kasnije odstranjivanje dio su normalnog razvoja mozga. Posebno je zanimljiva pojava prekobrojnih sinapsi u moždanoj kori u ranom postnatalnom razdoblju (između 8. mjeseca i 2. godine života); nakon tog razdoblja, broj sinapsi se smanjuje -isprva naglo, a potom sve sporije, pa odrasle vrijednosti dosegne tek nakon puberteta. To prekomjerno stvaranje i posljedično nestajanje prekobrojnih sinapsi, posebice u asocijacijskim područjima moždane kore, vezano je uz procese finog podešavanja asocijacijskih i komisurnih veza između neurona moždane kore, a odvija se najintenzivnije tijekom 2. godine života.
Moždana kora

Svi neuroni buduće moždane kore rađaju se u ventrikularnoj zoni telencefaličkog mjehurića i potom kroz intermedijalnu zonu migriraju prema pijalnoj površini i krajem 8. tjedna embrionalnog života oblikuju poseban gusti sloj, kortikalnu ploču, na granici intermedijalne i marginalne zone. Pojava kortikalne ploče znak je prijelaza iz embrionalnog u rano fetalno razdoblje. U ranom embrionalnom razdoblju stijenku telencefalona (kao i ostalih dijelova neuralne cijevi) oblikuju samo tri univerzalne embrionalne zone (VZ, IZ, MZ). Već krajem embrionalnog razdoblja (tijekom 6. tjedna trudnoće) pojavi se nova zona proliferacije karakteristična za telencefalon - subventrikularna zona (SVZ), smještena na granici VZ i IZ. Time već započinje regionalna histogeneza telencefalona. Potom, u bazolateralnom dijelu telencefaličkog mjehurića subventrikularna zona zadeblja i tako nastane ganglijski brežuljak, od kojeg se razviju bazalni gangliji telencefalona. U razdoblju između 12. i 15. tjedna trudnoće, razvije se nova subplate zona (SP) pa nakon toga stijenka telencefalona ima ukupno šest fetalnih zona: VZ, SVZ,IZ, SP, KP i MZ. Tri površnije zone (SP, KP iMZ) su razvojna osnova moždane kore. Iz fetalne intermedijalne zone se razvije bijela tvar odraslih moždanih polutki, a od proliferacijskih zona u odraslom mozgu preostane tek tanki sloj ependimskih stanica u stijenci moždanih komora.
Subplate zona i razvoj moždane kore

Opisani fetalni tip slojevite građe stijenke telencefalona najbolje je razvijen u razdoblju od 16. do 24. tjedna trudnoće. U ranijim razvojnim razdobljima prevladavaju procesi proliferacije i migracije, a u kasnijim razdobljima procesi diferencijacije neurona i sinaptogeneze. No, u ovom ključnom razdoblja istodobno se intenzivno odvijaju proliferacija, migracija, diferencijacija neurona, urastanje aksona i sinaptogeneza u osnovi moždane kore. Ključnu ulogu u tom razdoblju ima subplate zona jer ta zona ima zrele neurone što sintetiziraju različite neurotransmitere i neuropeptide, dok još neuroni za površinske dijelove kortikalne ploče tek nastaju u VZ i SVZ i migriraju kroz IZ; subplate zona je glavno mjesto rane sinaptogeneze i sadrži obilje sinapsi u ranom razdoblju, u kojem u kortikalnoj ploči još uopće nema sinapsi; subplate zona služi kao '''čekaonica'' za različite sustave aferentnih aksona što trebaju urasti u moždanu koru. Aferentni aksoni uspostave privremene sinapse na neuronima subplate zone i tu čekaju 2 do 3 mjeseca, a tek potom počnu urastati u samu kortikalnu ploču (između 24. i 28. tjedna trudnoće). U razdoblju najveće razvijenosti subplate zone (22. do 34. tjedan trudnoće), elementi kortikalnih neuronskih krugova imaju privremeni slojeviti raspored i privremena histokemijska i neurotransmiterska svojstva. Ti privremeni oblici organizacije tijekom kasnog fetalnog razdoblja su neurobiološka podloga privremenih oblika funkcionalne aktivnosti i obrazaca ponašanja u nedonoščadi pa je njihovo detaljno poznavanje važno za razumijevanje razvoja moždane kore čovjeka.
Tijekom tog razdoblja pojave se i prve regionalne i arealne razlike unutar moždane kore. Posljednji stadij prenatalnog života je razdoblje nezrele moždane kore novorođenčeta u kojoj se odvijaju intenzivni procesi histogenetske reorganizacije. U tom razdoblju započinje postupno nestajanje i/ili transformacija fetalnih slojeva i vrsta neurona, a odvija se opsežna histokemijska reorganizacija talamokortikalnog sustava. Ti se procesi nastavljaju se još dugo nakon rođenja. U ljudskoj moždanoj kori, "čekajući" asocijacijski i komisurni aksoni su glavni sastojak subplate zone nakon 28. tjedna trudnoće. Sljedeće važno obilježje kasnog fetalnog i perinatalnog razdoblja je pojava Brodmannovog temeljnog šestoslojnog tipa moždane kore (fetalni kortikalni slojevi I-VI) - no i ti slojevi podliježu značajnoj postnatalnoj reorganizaciji, pa se odrasli oblik Brodmannove kortikalne mape razvije nekoliko godina nakon rođenja.[5][6]
Razvoj bazalnih ganglija

Bazalni gangliji telencefalona nastaju od zadebljanog bazolateralnog dijela VZ i SVZ - od ganglijskog brežuljka. Tu se rađa i dio neurona talamusa, a vjerojatno i dio neurona moždane kore. Ganglijski brežuljak je tipična fetalna struktura što kasnije posve nestane. Od njegovog lateralnog dijela razviju se neuroni kaudatusa, putamena, klaustruma, amigdala i bazalne Meynertove jezgre, a od njegovog medijalnog dijela razviju se neuroni za nucleus accumbens septi (globus pallidus razvija se od diencefalona).
Osnovu strijatuma nalazimo već u 7. tjednu trudnoće, a pojavom unutarnje čahure sredinom 8. tjedna, osnova se podijeli u lateralni dio (osnova putamena) i manji medijalni dio (osnova kaudatusa). U 9. tjednu već se medijalno od putamena može uočiti i globus pallidus. Između 9. i 15. tjedna trudnoće, u osnovu strijatuma trajno pristižu novi neuroni i prva aferentna nigrostrijatalna vlakna. Oko 15. tjedna dolazi do dvije važne promjene: a) u dotad homogenom strijatumu pojave se prvi citoarhitektonski i histokemijski moduli i b) započinje intenzivna sinaptogeneza. Arhitektonski moduli između 17. i 24. tjedna trudnoće oblikuju složen mozaik, sastavljen od matriksa i u njega uklopljenih otočića, tzv. striosoma. Matriks i striosomi imaju različite neurotransmitere i različite neuronske veze. U kasnom fetalnom razdoblju (nakon 25. tjedna trudnoće) istodobno se dovršavaju svi glavni histogenetski procesi pa je to najbitnije razdoblje strukturnog razvoja strijatuma. Ganglijski brežuljak sadrži vrlo razvijenu mrežu proširenih venskih kapilara relativno debele stijenke. No, ta se kapilarna stijenka sastoji samo od endotela i tanke kolagene bazalne membrane što je posebno obilježje ventrikularne i subventrikularne zone ganglijskog brežuljka u usporedbi s okolnim tkivom.
Razvoj kralježnične moždine


Rostralni (prekordalni) dio neuralne cijevi preoblikuje se najprije u primarne, a zatim u sekundarne moždane mjehuriće, dok se od preostaloga (kaudalnog) dijela razvija kralježnična moždina. U dubini stijenke, uz šupljinu neuralne cijevi, dolazi do oblikovanja ventrikularne zone, gdje se stanice intenzivno množe. Od te proliferativne zone stanice se uspinju prema pijalnoj površini. Ventrikularna zona u području kralježnične moždine okružuje središnji kanal, canalis centralis. Središnji kanal i dalje komunicira s ostatkom šupljine neuralne cijevi. Još uvijek nediferencirane neuroektodermalne stanice mijenjaju oblik protežući tanke citoplazmatse izdanke u smjeru pijalne površine. Tijekom diobe tijela stanice se pomiču ponovno prema šupljini neuralne cijevi. Nakon završetka diobe neuroni, migrirajući prema vanjskoj strani, kao plašt oblikuju novi sloj, intermedijarnu zonu. Tijekom migracije neuroni pružaju nastavke koji prožimaju površnije smještenu marginalnu zonu, a sivu tvar čine tijela neurona u intermedijarnoj zoni. Nasuprot tome, izrastajući snopovi aksona nalaze se u marginalnoj zoni te oblikuju bijelu tvar. Zbog različitog rasta u debljinu, oblikuje se na unutarnjoj strani neuralne cijevi dugački uzdužni žlijeb, sulcus limitans, koji odjeljuje ventralno smještenu bazalnu ploču od dorzalno smještene krilne (alarne) ploče.

Bazalne ploče međusobno su povezane ventralnom pločom koja oblikuje dno središnjega kanala. Alarne su ploče međusobno povezane dorzalnom pločom koja čini pokrov središnjega kanala. Ventralna i dorzalna ploča zaostaju tijekom razvoja. Iz bazalne ploče razvijaju se veći ventralni i manji lateralni stupići jezgara kralježnične moždine. Ventralno se smještaju somatomotorni neuroni, a lateralno preganglionarni visceromotorni (vegetativni) neuroni. Tijela stanica alarne ploče oblikuju dorzalno stupić jezgara viscerosenzibilne te još dorzalnije somatosenzibilne (osjetne) neurone. Takvo grupiranje funkcionalno srodnih neurona u odijeljene stupiće jezgara zadržava se u odrasle kralježnične moždine.Konačni oblik kralježnična moždina poprima poradi proliferacije stanica u bazalnoj i alarnoj ploči, pa siva tvar dobiva oblik leptira, a zbog rasta aksona kroz marginalnu zonu uspostavljaju se veze s drugim dijelovima SŽS-a. Od šupljine neuralne cijevi preostaje ependimom okružen središnji kanal, canalis centralis, koji u odrasla čovjeka velikim dijelom obliterira.

Motoneuroni bazalne ploče koji kasnije oblikuju prednji rog kralježnične moždine, pružaju svoja eferentna vlakna u smjeru ciljnih organa, skeletnih mišića. Istodobno se od neuralnoga grebena odvajaju stanice. One se grupiraju s lateralne strane uzduž kralježnične moždine te kasnije diferenciraju u spinalne ganglije. U početku bipolarne stanice polako poprimaju tipičan oblik pseudounipolarnih neurona. Dendriti bipolarnih stanica uspostavljaju veze sa specifičnim receptorima ili kao slobodni živčani završeci odlaze u različite organe i kožu. Osjetne informacije kroz dendrite dolaze do tijela. Aksoni bipolarnih stanica kroz stražnji korijen ulaze u kralježničnu moždinu te završavaju na neuronima stražnjega roga ili se kroz stražnji snop (dorzalne kolumne) bez prekapčanja uzdižu do produžene moždine, medulla oblongata. Aferentni dendriti stanica spinalnih ganglija udružuju se u intervertebralnom otvoru, foramen intervertebrale, s eferentnim aksonima neurona prednjeg roga koji kroz prednji korijen napuštaju kralježničnu moždinu te oblikuju spinalne živce. Odsječak kralježnične moždine od kojeg se aksoni korjenova spajaju u jedan spinalni živac definira se kao odgovarajući segment kralježničke moždine (npr. segment L5 je odsječak kralježničke moždine čiji korijeni oblikuju 5. lumbalni spinalni živac).
U trećemu embrionalnomu mjesecu nalaze se odgovarajući segmenti kralježnične moždine i njima pridruženi intervertebralni otvori između dva tijela kralježaka na približno istoj visini. Daljnjim rastom tijela kralježnična moždina zaostaje u rastu prema koštanom dijelu kralježnice, te dolazi do uzdizanja kralježnične moždine u spinalnom kanalu, ascensus medullae spinalis. Na kraju petog mjeseca leži S1 segment kralježnične moždine približno 4 tijela više od prvog sakralnoga kralješka. Pri rođenju, vlakna koja napuštaju kralježnični kanal između S1 i S2 kralješka izlaze iz kralježnične moždine u visini L1 kralješka. U odraslih je kaudalni kraj kralježnične možidne, conus medullaris, otprilike u visini tijela L1 i L2 kralješka. Na toj se visini conus medullaris naglo suzuje u dio gdje nema neurona, filum terminale. On se hvata za kaudalni dio spinalnoga kanala te tako ostaje kao “trag” od ascensus medullae spinalis. Uz filum terminale izdužuju se aksoni u obliku konjskog repa kao cauda equina prema svojim intervertebralnim otvorima, gdje se ventralni i dorzalni korijeni ujedinjuju u spinalni živac.
Poremećaji razvoja
Poremećaj zatvaranja neuralne cijevi kod prednjega neuroporusa dovodi do pogrešnog oblikovanja mozga, a najdrastičniji primjer je anencefalija zbog koje dijete ne može preživjeti. Spina bifida je s druge strane poremećaj zatvaranja stražnjega neuroporusa, a može se pojaviti i u prikrivenom obliku, spina bifida occulta, gdje je došlo do nepotpunog zatvaranja moždinskoga kanala u kasnijemu tijeku razvoja, bez funkcionalnih ispada. Nasuprot tome, pri spina bifida aperta ne mogu se oblikovati kaudalni dijelovi kralježnične moždine. Ovisno o težini strukturnih promjena, može doći i do brojnih funkcionalnih poremećaja. Kod spine bifide totalis cjelokupna kralježnica (rachischisis) i kralježnična moždina (myeloschisis) su rascijepljene (nije spojivo sa životom). Kod spine bifide partialis kralježnica i kralježnična moždina mjestimično nisu zatvorene, a defekt može biti otvoren ili prekriven kožom s neurološkim ispadima (kljenuti, smetnje osjeta, smetnje pražnjenja mjehura). Kod Spine bifide occulte kralježnični lukovi iznad zdrave kralježnične moždine nisu srasli, koža je u pogođenu području obično jako dlakava; najčešće bez vidljivih neuroloških ispada.
Zbog nedostatka tkiva u središnjem dijelu mozga lateralne se možda komore spoje pa mozak ima jednu komoru, a očno polje se ne razdvoji što nazivamo holoprosencephalia i synophthalamia (spojene oči). Jedan od uzroka je mutacija gena sonic hedhehog (SHH) koji određuje središnju crtu središnjeg živčanog sustava na stadiju neuralne ploče.
 Anencefalija.
Anencefalija.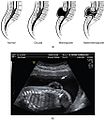 Spina bifida.
Spina bifida. Holoprosencephalia sa synophthalamiom.
Holoprosencephalia sa synophthalamiom.



Vidi još
Literatura i izvori
Vanjske poveznice
Wikiwand - on
Seamless Wikipedia browsing. On steroids.