Top Qs
Chronologie
Chat
Contexte
Ostracode
classe de crustacés De Wikipédia, l'encyclopédie libre

Remove ads
Les Ostracodes (Ostracoda) sont une classe de Crustacés microscopiques.
Ostracoda

Certaines espèces sont utilisées comme indicateurs de leur environnement actuel ou passé[1],[2]. Ils ont été testés comme bio-indicateurs de certaines pollutions, dont, en France, en baie de la Slack par exemple[3],[4].
Les estimations évaluent à plus de 20 000 le nombre d'espèces vivantes, dont seulement 8 000 ont été décrites[5]. Plus de 45 000 espèces et sous-espèces fossiles ont été décrites[6],[7],[8],[9].
Remove ads
Description




Les ostracodes possèdent une carapace bivalve faite de deux valves articulées dorsalement au niveau d'une structure appelée la charnière. Les valves ne portent pas de lignes de croissance. La carapace protège les parties molles et jusqu'à huit paires d'appendices chez les adultes, ainsi que des appendices copulatoires et une furca. Seules les extrémités de quelques appendices sortent par l'espace entre les valves ventralement quand les animaux se déplacent sur le substrat, se nourrissent ou quand ils nagent dans la colonne d'eau. Les ostracodes ont des juvéniles, pas des larves, et se développent par mues successives ; en général, les adultes ne muent pas[10].
Leur taille varie de 0,3 mm à environ 5 cm (0,5–3 mm pour la plupart des espèces).
Remove ads
Habitat, répartition
La plupart des espèces d'ostracodes vivent dans les environnements aquatiques, ils sont généralement benthiques (ou necto-benthiques) et interstitiels[11],[12],[13]. Ils sont communément trouvés dans la plupart des habitats aquatiques, des grands fonds marins aux mares temporaires, rivières, estuaires, sources chaudes. Les espèces planctoniques sont communes dans les habitats marins mais pas dans les masses d'eaux continentales[14].
Certains groupes d'ostracodes, notamment certains membres des Cypridoidea (Scotiinae et Candoninae), Cytheroidea, Darwinuloidea et Terrestricytheroidea, peuvent même peupler les environnements semi-terrestres[15]. D'autres sont épibiontes sur d'autres crustacés, c'est le cas des Entocytheridae[16].
La plupart des ostracodes marins sont sténohalins, mais il existe une grande diversité d'espèces plus ou moins euryhalines qui occupent des environnements côtiers à la salinité fluctuantes[11],[12]. Les espèces Cyprideis torosa et Eucypris mareotica sont probablement parmi les plus halotolérantes, trouvées respectivement à des salinités jusqu'à 180g/L et 110 g/l[17],[18].
Remove ads
Reproduction
La plupart des espèces d'ostracodes vivant dans les milieux marins et les lacs anciens ont une reproduction sexuée, tandis que la plupart des formes d'eau douce ont une reproduction asexuée ou mixte[19],[20],[21]. La sexualité présente des avantages par rapport à l'asexualité dans des environnements géologiquement stables mais écologiquement instables[20]. Selon les espèces, les œufs peuvent être pondus ou couvés[19],[22].
Histoire évolutive
La carapace calcaire des ostracodes leur a permis d'avoir les archives fossiles les plus complètes des arthropodes, s'étendant sur environ 450 millions d'années (Ma) depuis l'Ordovicien[23],[24]. Leur présence dans les milieux marins au Cambrien a longtemps été discutée (Bradoriida et Phosphatocopida) mais la découverte de spécimens aux parties molles conservées par phosphatisation a montré qu'ils ne sont pas des ostracodes[25]. Les plus anciens ostracodes connus datent du Trémadocien, à l'Ordovicien inférieur, biozone à conodonte Paltodus deltifer[25]. Des analyses moléculaires suggèrent que les ostracodes se placent à la place ou proche de la base des Pancrustacés suggérant une origine possible à la fin du Protérozoïque[26],[27]. Des mandibules phosphatisées dans des dépôts du Cambrien supérieur de l'ouest du Canada à la morphologie proche des ostracodes seraient les premières preuves formelles de l'existence d'ostracodes avant l'Ordovicien[28].
Les ostracodes sont un modèle unique de l'évolution du sexe des métazoaires : un ostracode de l'espèce Colymbosathon ecplecticos du Silurien est le plus ancien métazoaire mâle connu[29].
Remove ads
Spermatozoïdes fossiles
Résumé
Contexte
Des spermatozoïdes d'ostracodes fossiles vieux de 17 millions d'années ont été découverts[30]. Les recherches, objet d'une coopération internationale autour d'une équipe de l'université de Nouvelle-Galles du Sud, ont été menées à partir de 66 échantillons trouvés en 1988 sur le site fossilifère de Riversleigh, dans l’État du Queensland (Australie). Elles ont mis en évidence la présence de la structure hélicoïdale typique des organites à l'aide desquelles les spermatozoïdes des ostracodes se déplacent. La membrane nucléaire de ces gamètes est restée visible, bien que l'ADN qu'elle contenait ait disparu. Bien que difficiles à mesurer avec précision, ce sont, comme c'est classiquement le cas pour ces espèces, des spermatozoïdes géants, d'une taille du même ordre de grandeur que celle du corps de l'animal (de 1,2 à 1,3 mm). Les mâles de certaines espèces produisent des spermatozoïdes d'une longueur de 6 mm, soit dix fois leur taille[31]. Sur les femelles ont été relevés des conduits internes d'une longueur pouvant atteindre quatre fois celle du corps de l'animal, également comparable à ce qui peut être observé sur les espèces actuelles. L'ensemble des observations indique un mode de reproduction resté inchangé depuis 17 millions d'années[31]. Le sperme, sans atteindre l'âge de celui d'un spécimen de collembole pris dans l'ambre il y a 40 millions d'années, devient cependant, au moment de sa découverte, le plus ancien jamais trouvé dans un spécimen fossilisé. Riches en phosphore, les fientes de chauves-souris dont le repaire surplombait la mare où vivaient les ostracodes, ont pu jouer un rôle dans la préservation de leurs tissus mous[32].
Remove ads
Bioluminescence
Certains ostracodes ont un organe bio-luminescent qu'ils utilisent comme défense contre la prédation[31] et, dans quelques cas seulement présents dans les Caraïbes, pour l'accouplement[33].
Liste des sous-taxons
Taxons actuels
- sous-classe Myodocopa Sars, 1866
- ordre Myodocopida Sars, 1866
- famille Cypridinidae Baird, 1850
- sous-famille Cypridininae Baird, 1850
- genre Cypridina Milne-Edwards, 1840
- sous-famille Cypridininae Baird, 1850
- famille Cypridinidae Baird, 1850
- ordre Halocyprida
- ordre Myodocopida Sars, 1866
- sous-classe Podocopa Sars, 1866
- ordre Platycopida
- ordre Podocopida
- ordre Palaeocopida †
Selon World Register of Marine Species (10 mars 2017)[34] :
- sous-classe Archaeocopa †
- sous-classe Bradoriida †
- sous-classe Metacopa †
- sous-classe Myodocopa
- sous-classe Palaeocopa
- sous-classe Platycopa
- sous-classe Podocopa
- Ostracoda incertae sedis
Taxons fossiles
- ordre †Kloedenellocopida
- ordre †Palaeocopida
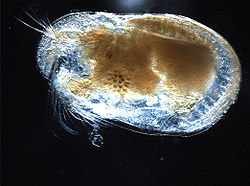 Un ostracode de la famille des Cylindroleberididae (ordre des Myodocopida)
Un ostracode de la famille des Cylindroleberididae (ordre des Myodocopida)

Remove ads
Notes et références
Voir aussi
Wikiwand - on
Seamless Wikipedia browsing. On steroids.
Remove ads