Silurien
troisième période géologique du Paléozoïque, 443,4 à 419,2 Ma avant le présent De Wikipédia, l'encyclopédie libre

Le Silurien, parfois anciennement nommé Gothlandien, est un système géologique qui s'étend de −443,4 à −419,2 Ma. La datation de début et de fin, bien que définie avec précision par les couches stratigraphiques de référence, est connue à seulement quelques millions d'années près. Il est précédé de l'Ordovicien et suivi du Dévonien. La fin de l'Ordovicien est marquée par l'une des cinq grandes extinctions massives de l'histoire des temps géologiques, où près de 60 % des espèces marines ont disparu et peut-être 85 % du total des espèces (végétales et animales) : l'extinction Ordovicien-Silurien.
Silurien
Gothlandien, Silurien s.s.
Gothlandien, Silurien s.s.
| Notation chronostratigraphique | S |
|---|---|
| Notation française | s |
| Notation RGF | s |
| Équivalences | Le Silurien s.l. inclut l'Ordovicien |
| Niveau | Période / Système |
| Érathème / Ère - Éonothème / Éon |
Paléozoïque Phanérozoïque |
Stratigraphie
| Début | Fin |
|---|---|
| 443,8 ± 1,5 Ma (Extinction Ordovicien-Silurien) |
419,2 ± 3,2 Ma |
Paléogéographie et climat

Reconstitution de la géographie terrestre du Silurien (440 Ma)
| Taux de O2 atmosphérique |
env. 14 %vol[1] (70 % de l'actuel) |
|---|---|
| Taux de CO2 atmosphérique |
env. 4 500 ppm[2] (16 fois le niveau d'avant la révolution industrielle) |
| Température moyenne |
17 °C[3] (+3 °C par rapport à l'actuel) |
| Niveau moyen des mers | +180 m (par rapport à l'actuel)[4] |
- fin de l'orogenèse calédonienne
- 440 Ma : formation du sol par les algues
Faune et flore

- 440 Ma : sortie de l'eau des algues
- 435 Ma : arachnides
- 430 Ma : gnathostomes (vertébrés à mâchoire)
- 430 Ma : appendices pairs antérieurs des vertébrés
- 428 Ma : myriapodes
- 420 Ma : ostéichtyens (vertébrés à squelette minéralisé)
- 420 Ma : actinoptérygiens (poissons à nageoires rayonnés)
- 420 Ma : sortie de l'eau des plantes vasculaires
Le Silurien a été décrit par Roderick Murchison en 1839 (The Silurian System) se basant sur des terrains découverts dans le sud du Pays de Galles. Il a été nommé en 1835 d'après une tribu celtique : les Silures. Cette classification est entrée en conflit avec celle d'Adam Sedgwick du Cambrien ; ce problème a été résolu par Charles Lapworth en ajoutant l'Ordovicien entre le Cambrien et le Silurien pour les couches géologiques contestées. Le nom « Gothlandien » provient des gisements de l'île suédoise de Gotland.
Subdivisions
Le Silurien est divisé en quatre séries ou époques, bien que certains auteurs le découpaient autrefois en trois (Silurien supérieur, moyen et inférieur). Les datations des subdivisions ont été revues par la Commission internationale de stratigraphie (ICS) en 2012[5].
| Pridolien | (423,0 ± 2,3 – 419,2 ± 3,2 Ma) |
| Ludlowien | |
| Ludfordien | (425,6 ± 0,9 – 423,0 ± 2,3 Ma) |
| Gorstien | (427,4 ± 0,5 – 425,6 ± 0,9 Ma) |
| Wenlockien | |
| Homérien | (430,5 ± 0,7 – 427,4 ± 0,5 Ma) |
| Sheinwoodien | (433,4 ± 0,8 – 430,5 ± 0,7 Ma) |
| Llandoveryen | |
| Télychien | (438,5 ± 1,1 – 433,4 ± 0,8 Ma) |
| Aéronien | (440,8 ± 1,2 – 438,5 ± 1,1 Ma) |
| Rhuddanien | (443,4 ± 1,5 – 440,8 ± 1,2 Ma) |
Paléogéographie
Pendant le Silurien, le Gondwana reste dans les latitudes hautes de l'hémisphère sud mais la calotte glaciaire est moins étendue que pendant l'Ordovicien. Les autres continents se rapprochent pour commencer la formation d'un second supercontinent : la Laurussia, ce qui ferme l'océan Iapetus qui séparait les divers continents de l'hémisphère nord avant leur accrétion. Ceci a donné naissance à l'orogenèse calédonienne dont on retrouve les traces en Scandinavie. Cette grande chaine de montagnes se poursuit vers l'actuel sud-ouest vers le Canada et les États-Unis où elle forme les Appalaches.
Les petits terranes accrétés au Gondwana pendant l'Ordovicien se détachent à nouveau pour former le superterrane hunnique, appelé ainsi car constitué de la majeure partie des territoires dévastés par Attila[6]. Des couches d'hématite — un minerai de fer — ont été importantes pour l'économie en Amérique du Nord.
Faune silurienne
La vie est abondante notamment en milieu marin, car le niveau des océans est élevé en Europe et dans le nord-est de l'Amérique. Toutefois des plantes multicellulaires encore simples, à spores, colonisent les terres, accompagnées par des arthropodes. Les poissons se sont diversifiés considérablement et ont développé des écailles mobiles. Une faune diverse de scorpions de mer, certains longs de plusieurs mètres, se trouve en Amérique du Nord. Échinodermes, mollusques et graptolites sont toujours abondants.
Le Silurien en France
À la base des schistes verts de l'unité de Génis, dans le Bas Limousin, apparaissent des lentilles d'un calcaire à entroques contenant des conodontes du Silurien supérieur[7].
Un transect géologique au niveau de la région du houtland en Flandre montre un socle de schistes datant du Silurien vers 300–400 m de profondeur[8].
Climat et niveau de la mer
Résumé
Contexte
On pensait autrefois que la période du Silurien avait bénéficié de températures relativement stables et chaudes, contrairement aux glaciations extrêmes de l'Ordovicien qui l'avaient précédé et à la chaleur extrême du Dévonien qui a suivi ; cependant, on sait maintenant que le climat mondial a subi de nombreuses fluctuations drastiques tout au long du Silurien[9],[10], comme en témoignent de nombreuses excursions majeures des isotopes du carbone et de l'oxygène au cours de cette période géologique[11],[12],[13]. Le niveau de la mer s'est élevé à partir de son minimum hirnantien tout au long de la première moitié du Silurien ; ils ont ensuite diminué pendant le reste de la période, même si des tendances à plus petite échelle se superposent à cette tendance générale ; quinze hauts plateaux (périodes pendant lesquelles le niveau de la mer était au-dessus du bord du plateau continental) peuvent être identifiés, et le niveau de la mer le plus élevé du Silurien était probablement d'environ 140 mètres plus haut que le niveau le plus bas atteint[14].
Au cours de cette période, la Terre est entrée dans une phase de serre chaude, soutenue par des niveaux élevés de CO2 de 4 500 ppm, et des mers chaudes et peu profondes couvraient une grande partie des masses terrestres équatoriales[15]. Au début du Silurien, les glaciers se sont retirés vers le pôle Sud jusqu'à ce qu'ils disparaissent presque au milieu du Silurien[10]. Les couches de coquilles brisées (appelées coquina) fournissent des preuves solides d'un climat dominé par de violentes tempêtes générées alors comme aujourd'hui par les surfaces marines chaudes[16].
Perturbations
Le climat et le cycle du carbone semblent plutôt instables au cours du Silurien, qui présentait une fréquence d'excursions isotopiques (indicatrice de fluctuations climatiques) plus élevée que toute autre période[14]. L’événement Ireviken, l’événement Mulde et l’événement Lau représentent chacun des excursions isotopiques consécutives à une extinction massive mineure[17] et associées à un changement rapide du niveau de la mer. Chacun laisse une signature similaire dans les archives géologiques, à la fois géochimiques et biologiques ; les organismes pélagiques (nageant librement) ont été particulièrement durement touchés, tout comme les brachiopodes, les coraux et les trilobites, et les extinctions se produisent rarement dans une série rapide d'éclats rapides[14],[13]. Les fluctuations climatiques s'expliquent mieux par une séquence de glaciations, mais le manque de tillites du Silurien moyen à supérieur rend cette explication problématique[18].
Flore et faune
Résumé
Contexte
La période silurienne a été considérée par certains paléontologues comme un intervalle de récupération prolongé après l'extinction massive de l'Ordovicien supérieur (LOME, pour l'anglais Late Ordovician Mass Extinction), qui a interrompu l'augmentation en cascade de la biodiversité qui s'était continuellement poursuivie tout au long du Cambrien et de la majeure partie de l'Ordovicien[19].
Le Silurien a été la première période à voir des mégafossiles d'un vaste biote terrestre sous la forme de forêts miniatures ressemblant à de la mousse le long des lacs et des ruisseaux et des réseaux de grands nématophytes mycorhiziens, annonçant le début de la révolution terrestre silurienne-dévonienne[20],[21],[22]. Cependant, la faune terrestre n'a pas eu d'impact majeur sur la Terre jusqu'à ce qu'elle se diversifie au Dévonien[14].
Les premières traces fossiles de plantes vasculaires, c'est-à-dire de plantes terrestres dont les tissus transportent l'eau et la nourriture, sont apparues dans la seconde moitié de la période silurienne[23]. Les premiers représentants connus de ce groupe sont Cooksonia. La plupart des sédiments contenant Cooksonia sont de nature marine. Les habitats préférés se trouvaient probablement le long des rivières et des ruisseaux. Baragwanathia semble être presque aussi vieux, datant du début de Ludlow (420 millions d'années) et a des tiges ramifiées et des feuilles en forme d'aiguilles de 10 à 20 centimètres. La plante présente un haut degré de développement par rapport à l'âge de ses restes fossiles. Des fossiles de cette plante ont été enregistrés en Australie[24],[25] au Canada[26] et en Chine[27]. Eohostimella heathana est une « plante » précoce, probablement terrestre, connue à partir de fossiles de compression[28] de l'âge du Silurien inférieur (Llandovery)[29].[43] La chimie de ses fossiles est similaire à celle des plantes vasculaires fossilisées, plutôt qu'à celle des algues[28].
Des fossiles considérés comme des animaux terrestres sont également connus du Silurien. Le plus ancien signalement définitif de mille-pattes jamais connu est Kampecaris obanensis et Archidesmus sp. du Silurien supérieur (il y a 425 millions d'années) de Kerrera[30]. Il existe également d'autres mille-pattes, mille-pattes et arachnoïdes trigonotarbidés connus à Ludlow (il y a 420 millions d'années)[30],[31],[32]. Les invertébrés prédateurs indiqueraient qu’il existe des réseaux trophiques simples comprenant des proies non prédatrices. En extrapolant à partir du biote du Dévonien inférieur, Andrew Jeram et al. en 1990[33] a suggéré un réseau trophique basé sur des détritivores et des brouteurs de micro-organismes encore inconnus[34]. Les mille-pattes de la formation de Cowie tels que Cowiedesmus et Pneumodesmus étaient considérés comme les mille-pattes les plus anciens du Silurien moyen il y a 428 millions d'années[35],[36]. Bien que l'âge de cette formation soit plus tard réinterprété comme étant celui du Dévonien inférieur par certains chercheurs[37], une étude plus ultérieure en 2023 a reconfirmé son âge comme étant la fin de l'époque Wenlock du Silurien moyen[38]. Quoi qu'il en soit, Pneumodesmus est toujours un fossile important en tant que preuve définitive la plus ancienne de stigmates permettant de respirer dans l'air.
Apparurent les premiers poissons osseux, les Osteichthyes, représentés par les Acanthodiens couverts d'écailles osseuses. Les poissons ont atteint une diversité considérable et ont développé des mâchoires mobiles, adaptées des supports des deux ou trois arcs branchiaux antérieurs. Une faune diversifiée d'euryptérides (scorpions marins), certains mesurant plusieurs mètres de long, rôdait dans les mers et les lacs peu profonds du Silurien d'Amérique du Nord ; bon nombre de leurs fossiles ont été découverts dans l’État de New York. Les sangsues ont également fait leur apparition au cours de la période silurienne. Les brachiopodes étaient abondants et diversifiés, la composition taxonomique, l'écologie et la biodiversité des brachiopodes du Silurien reflétant celles de l'Ordovicien[39]. Les brachiopodes qui ont survécu à LOME ont développé de nouvelles adaptations au stress environnemental[40] et ont eu tendance à être endémiques à une seule paléoplaque au lendemain de l'extinction massive, mais ont ensuite élargi leur aire de répartition[41]. Les brachiopodes les plus abondants étaient les atrypides et les pentamrides[42] ; les atrypides ont été les premiers à se rétablir et à se rediversifier dans le Rhuddanien après LOME[43], tandis que la récupération des pentamérides a été retardée jusqu'à l'Aéronien[42]. Les bryozoaires présentaient des degrés significatifs d'endémisme sur un plateau particulier[44]. Ils ont également développé des relations symbiotiques avec les cnidaires[45] et les stromatolites[46]. De nombreux fossiles de bivalves ont également été trouvés dans les gisements du Silurien[47] et les premiers bivalves profonds sont connus de cette période[48]. Les chitons ont connu un pic de diversité au milieu du Silurien[49]. Les hédérelloïdes ont connu un succès significatif au Silurien, avec quelques symbioses développantes avec le corail rugueux colonial Entelophyllum[50]. Le Silurien a été l'apogée des tentaculitoïdes[51], qui ont connu un rayonnement évolutif concentré principalement en Baltoscandie[52], ainsi qu'une expansion de leur aire de répartition géographique dans le Llandovery et le Wenlock[53]. Les trilobites ont commencé à se rétablir au Rhuddanien[54] et ont continué à connaître du succès au Silurien comme ils l'avaient fait à l'Ordovicien malgré la réduction de la diversité des clades à la suite de LOME[55]. Le Silurien inférieur a été une période chaotique de renouvellement pour les crinoïdes alors qu'ils se rediverversaient après LOME[56]. Les membres de Flexibilia, qui ont été peu impactés par LOME, ont pris une importance écologique croissante dans les mers du Silurien[57]. Les camerates monobathridés, comme les flexibles, se sont diversifiés dans le Llandovery, tandis que les cyathocrinidés et les dendrocrinidés se sont diversifiés plus tard au Silurien[58]. Les lobolithes scyphocrinoïdes sont soudainement apparus dans le Silurien terminal, peu avant la limite Silurien-Dévonien, et ont disparu aussi brusquement qu'ils sont apparus très peu de temps après leur première apparition[59]. Les symbiotes endobiotiques étaient courants dans les coraux et les stromatoporoïdes[60],[61]. Les coraux rugueux en particulier étaient colonisés et incrustés par une gamme diversifiée d'épibiontes[62], y compris certains hédérelloïdes comme mentionné ci-dessus[50]. Les scléractiniens photosymbiotiques ont fait leur première apparition au Silurien moyen[63]. L'abondance des récifs était inégale ; parfois, les fossiles sont fréquents, mais à d'autres endroits, ils sont pratiquement absents des archives rocheuses[14].
 Cooksonia, la première plante vasculaire, Silurien moyen
Cooksonia, la première plante vasculaire, Silurien moyen Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK
Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK Fragments de crinoïdes dans un calcaire du Silurien (Pridoli) (Saaremaa, Estonie)
Fragments de crinoïdes dans un calcaire du Silurien (Pridoli) (Saaremaa, Estonie) Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK
Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK Eurypterus, un euryptéride commun du Silurien supérieur
Eurypterus, un euryptéride commun du Silurien supérieur Pterygotus était un eurypteridé géant qui avait une distribution presque cosmopolite (la reconstruction présentée ici est l'espèce dévonienne P. anglicus)
Pterygotus était un eurypteridé géant qui avait une distribution presque cosmopolite (la reconstruction présentée ici est l'espèce dévonienne P. anglicus) Les trilobites étaient encore divers et communs au Silurien. Les fossiles de Calymene celebra sont extrêmement abondants dans certaines parties du centre des États-Unis.
Les trilobites étaient encore divers et communs au Silurien. Les fossiles de Calymene celebra sont extrêmement abondants dans certaines parties du centre des États-Unis. Halysites étaient un corail tabulé, un groupe éteint qui a vécu au Paléozoïque.
Halysites étaient un corail tabulé, un groupe éteint qui a vécu au Paléozoïque. Parioscorpio était un arthropode énigmatique du Silurien du Wisconsin.
Parioscorpio était un arthropode énigmatique du Silurien du Wisconsin. Un spécimen de Dalmanites limulurus Dalmanites limulurus provenant des strates siluriennes de New York
Un spécimen de Dalmanites limulurus Dalmanites limulurus provenant des strates siluriennes de New York Une roche contenant plusieurs brachiopodes pentaméridés géodisés provenant de strates de l'Indiana.
Une roche contenant plusieurs brachiopodes pentaméridés géodisés provenant de strates de l'Indiana.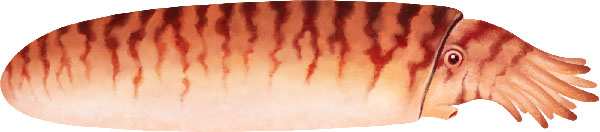 Sphooceras était un céphalopode nautiloïde trouvé dans les strates siluriennes de la République tchèque
Sphooceras était un céphalopode nautiloïde trouvé dans les strates siluriennes de la République tchèque Jamoytius était un vertébré énigmatique qui pourrait être apparenté au poisson Anaspid.
Jamoytius était un vertébré énigmatique qui pourrait être apparenté au poisson Anaspid.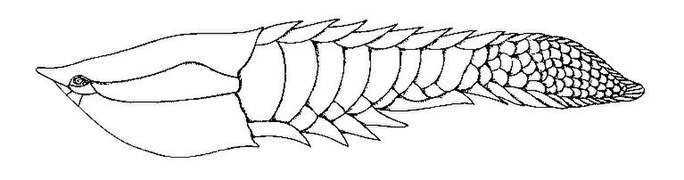 Poraspis, un genre de poisson blindé sans mâchoire du Silurien supérieur du Canada, de la Norvège et des États-Unis.
Poraspis, un genre de poisson blindé sans mâchoire du Silurien supérieur du Canada, de la Norvège et des États-Unis. Tujiaaspis est un agnathan galeaspid du début du Silurien (Telychian) de Chine, montrant l'origine de nageoires appariées
Tujiaaspis est un agnathan galeaspid du début du Silurien (Telychian) de Chine, montrant l'origine de nageoires appariées Qianodus[64] est un genre de chondrichtyen à base de dents du début du Silurien (Aéronien) de Chine.
Qianodus[64] est un genre de chondrichtyen à base de dents du début du Silurien (Aéronien) de Chine. Fanjingshania, requin épineux climatiidé du Silurien inférieur (Aéronien) décrit à partir d'éléments dermosquelettiques désarticulés
Fanjingshania, requin épineux climatiidé du Silurien inférieur (Aéronien) décrit à partir d'éléments dermosquelettiques désarticulés Shenacanthus[65] est un genre de Chondrichtyen à tige mâchoire du Silurien inférieur (Telychien) de Chine.
Shenacanthus[65] est un genre de Chondrichtyen à tige mâchoire du Silurien inférieur (Telychien) de Chine. Xiushanosteus[65] is the oldest known placoderm from the early Silurian (Telychian) of China
Xiushanosteus[65] is the oldest known placoderm from the early Silurian (Telychian) of China Entelognathus primordialis est le plus ancien placoderme connu du début du Silurien (Telychien) de Chine.
Entelognathus primordialis est le plus ancien placoderme connu du début du Silurien (Telychien) de Chine.










_1.jpg)









Voir aussi
Articles connexes
Liens externes
Notes et références
Wikiwand - on
Seamless Wikipedia browsing. On steroids.